

The adhesion GPCRs CELSR1-3 and LPHN3 engage G proteins via distinct activation mechanisms
- PMID: 37224017
- PMCID: PMC10592476
- DOI: 10.1016/j.celrep.2023.112552
The adhesion GPCRs CELSR1-3 and LPHN3 engage G proteins via distinct activation mechanisms
Abstract
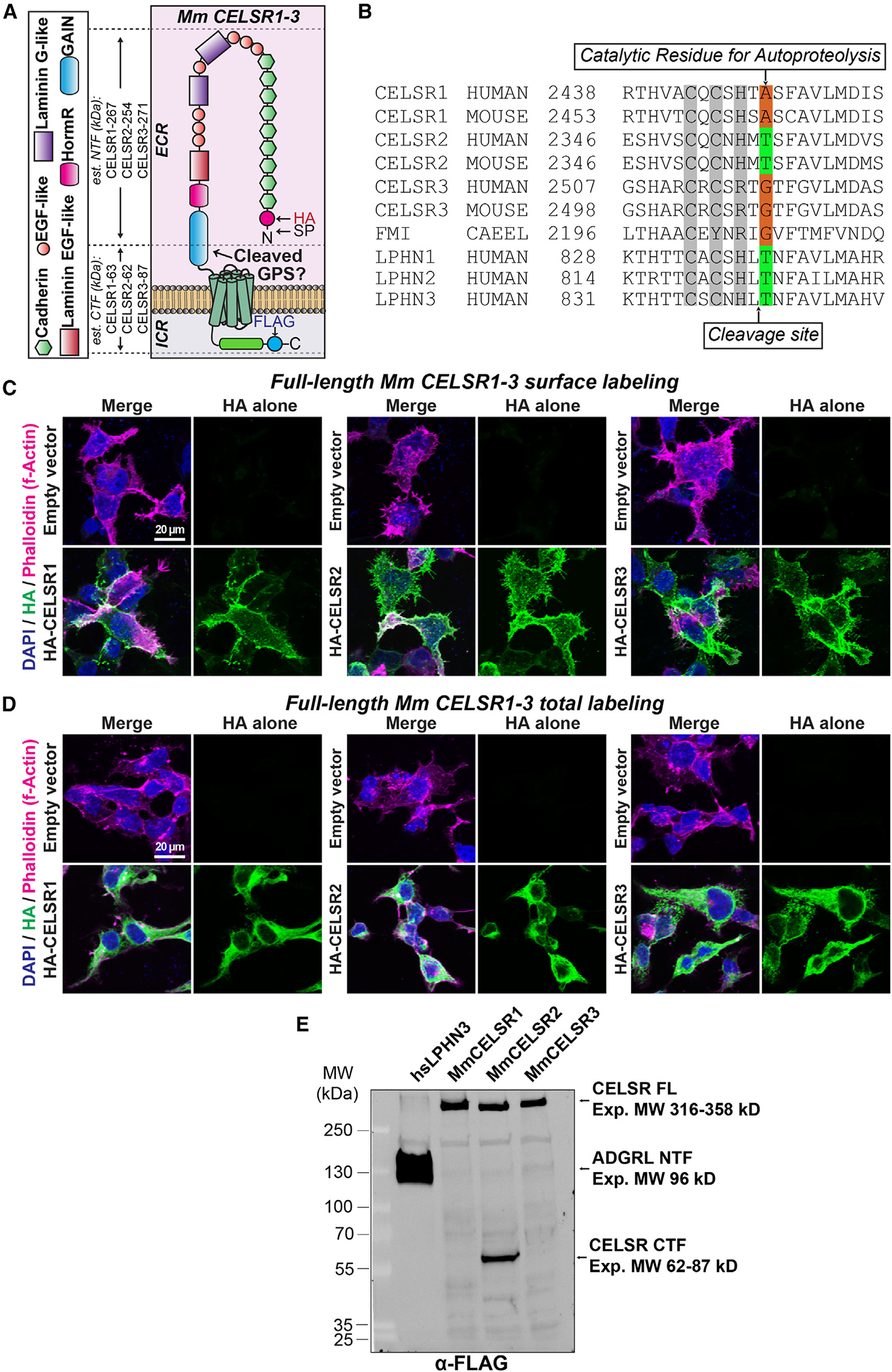
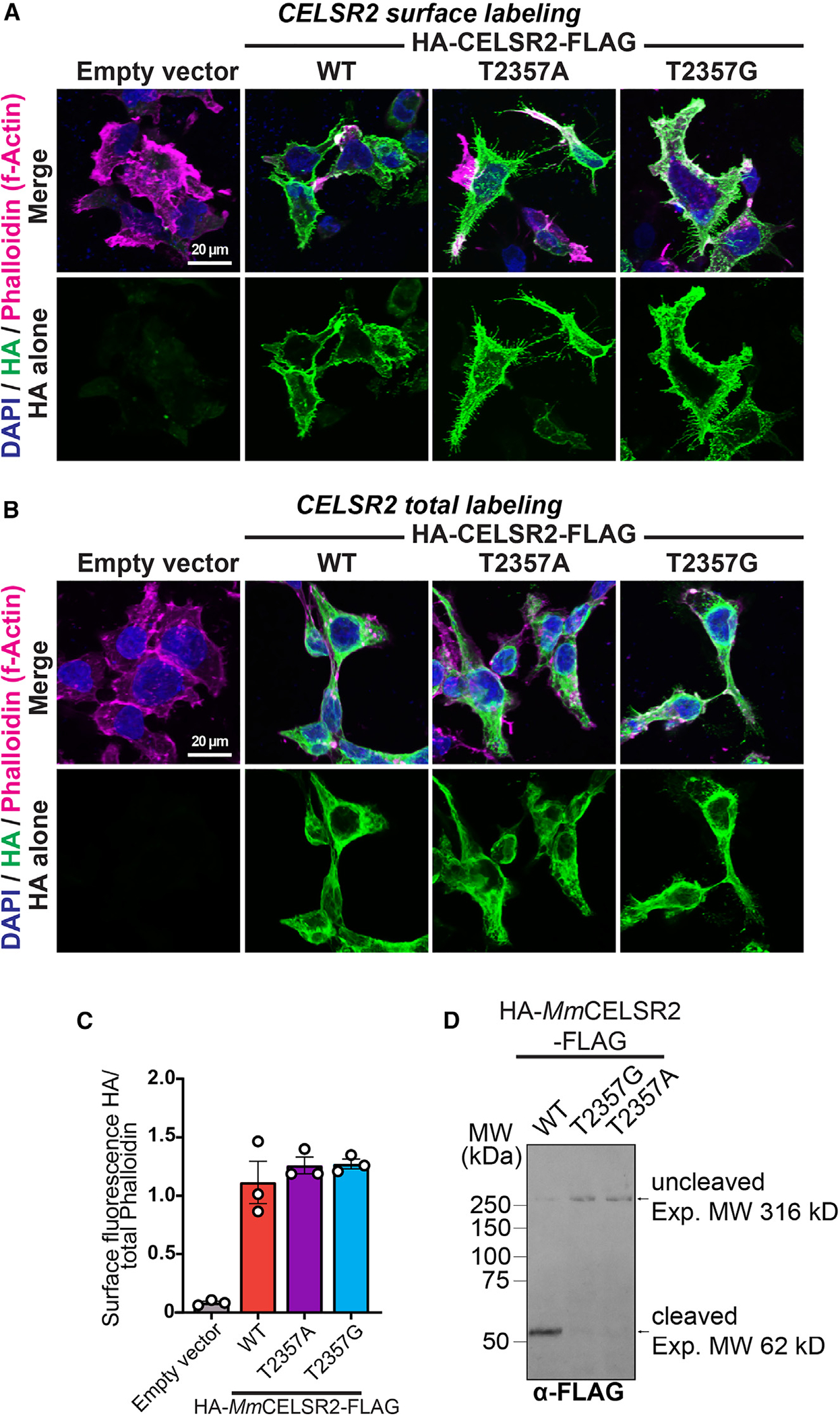
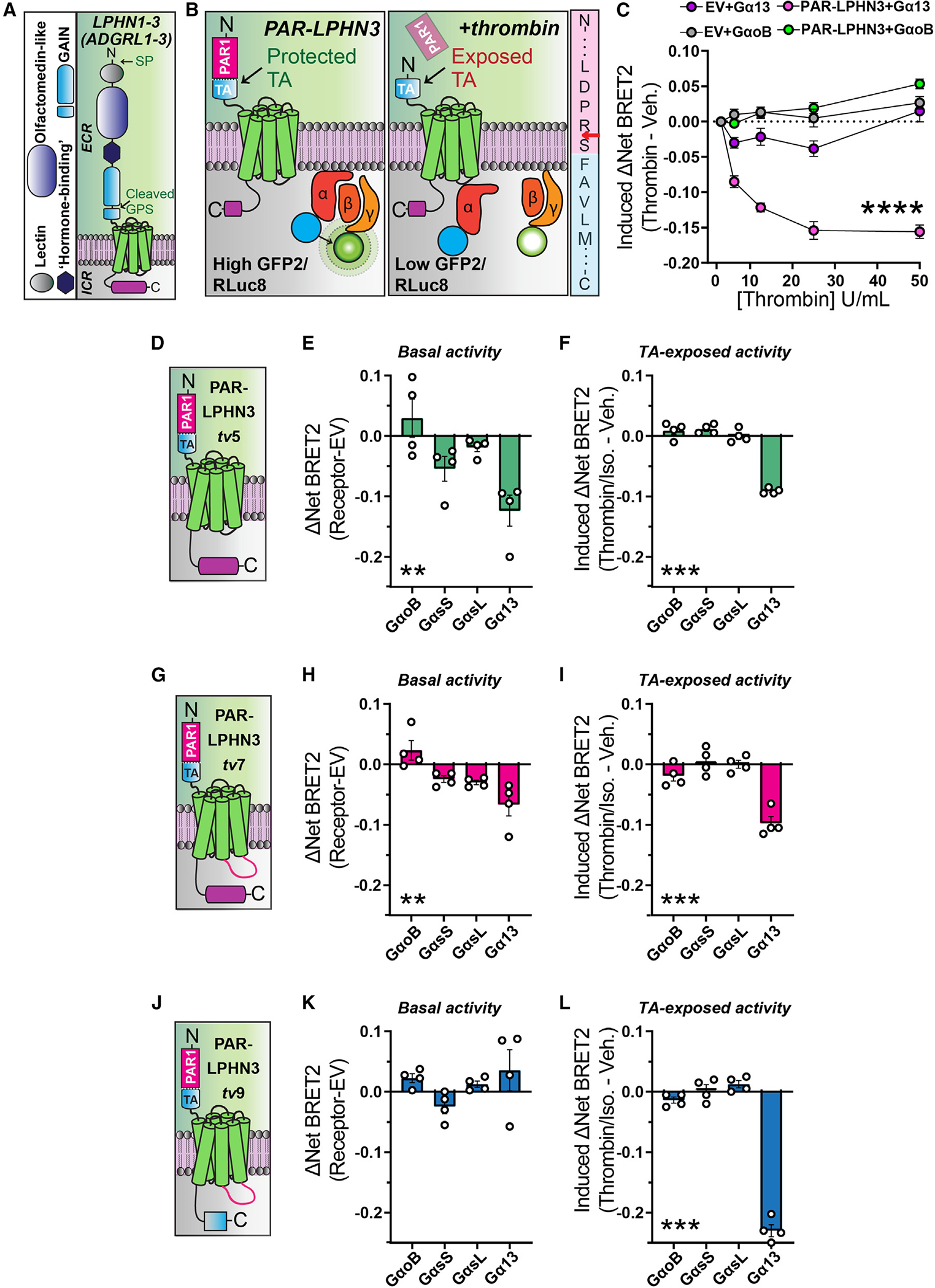
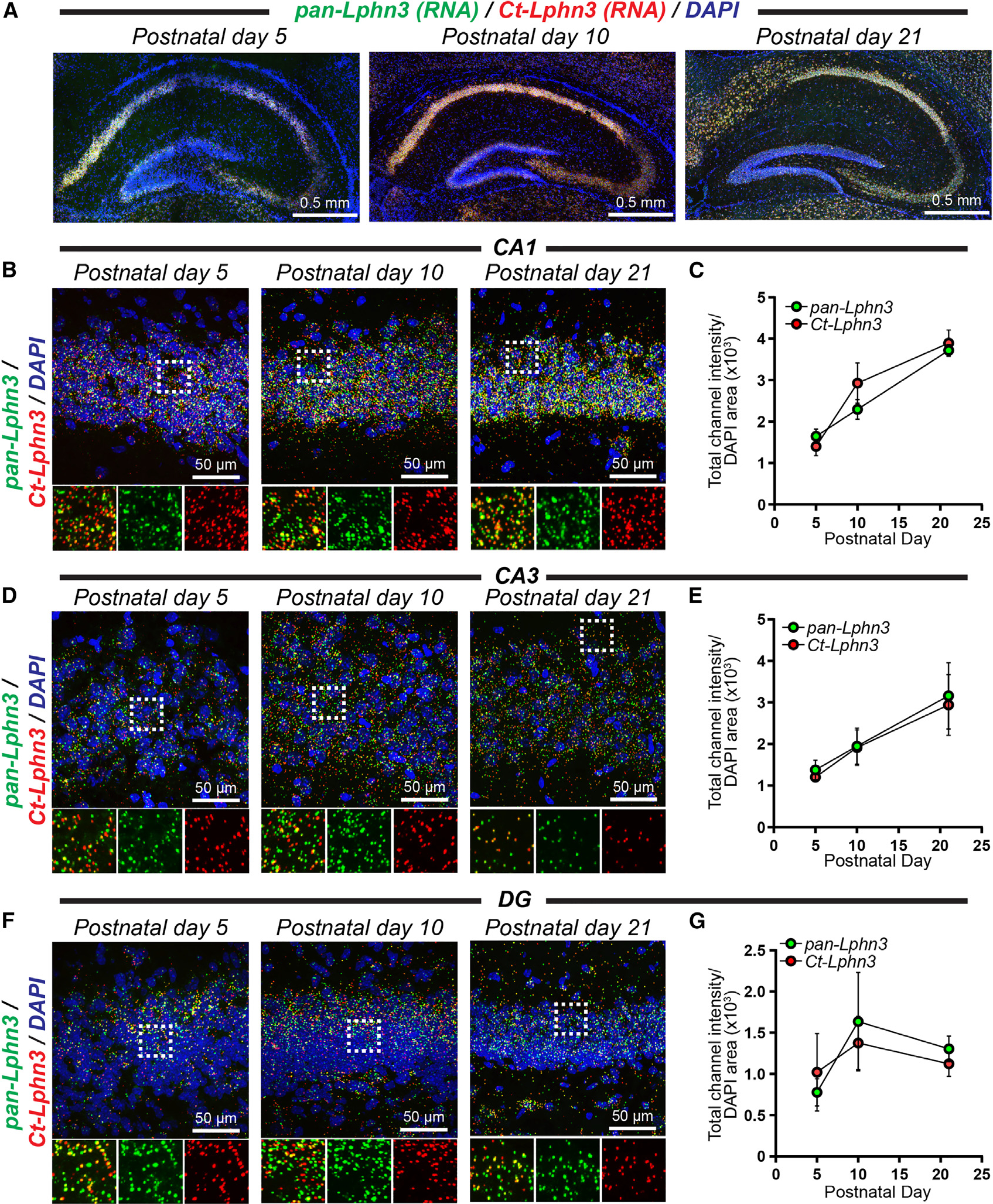
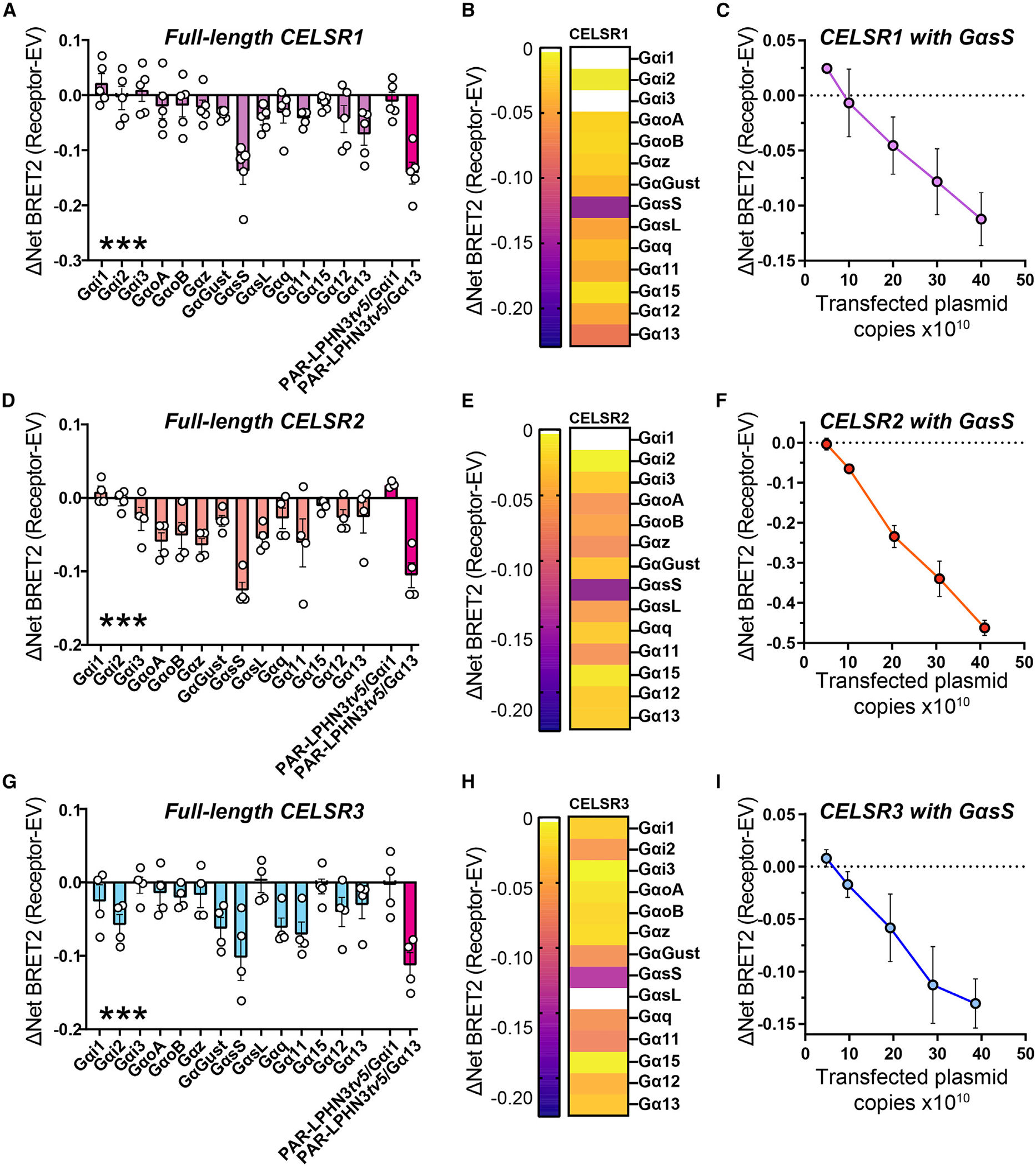
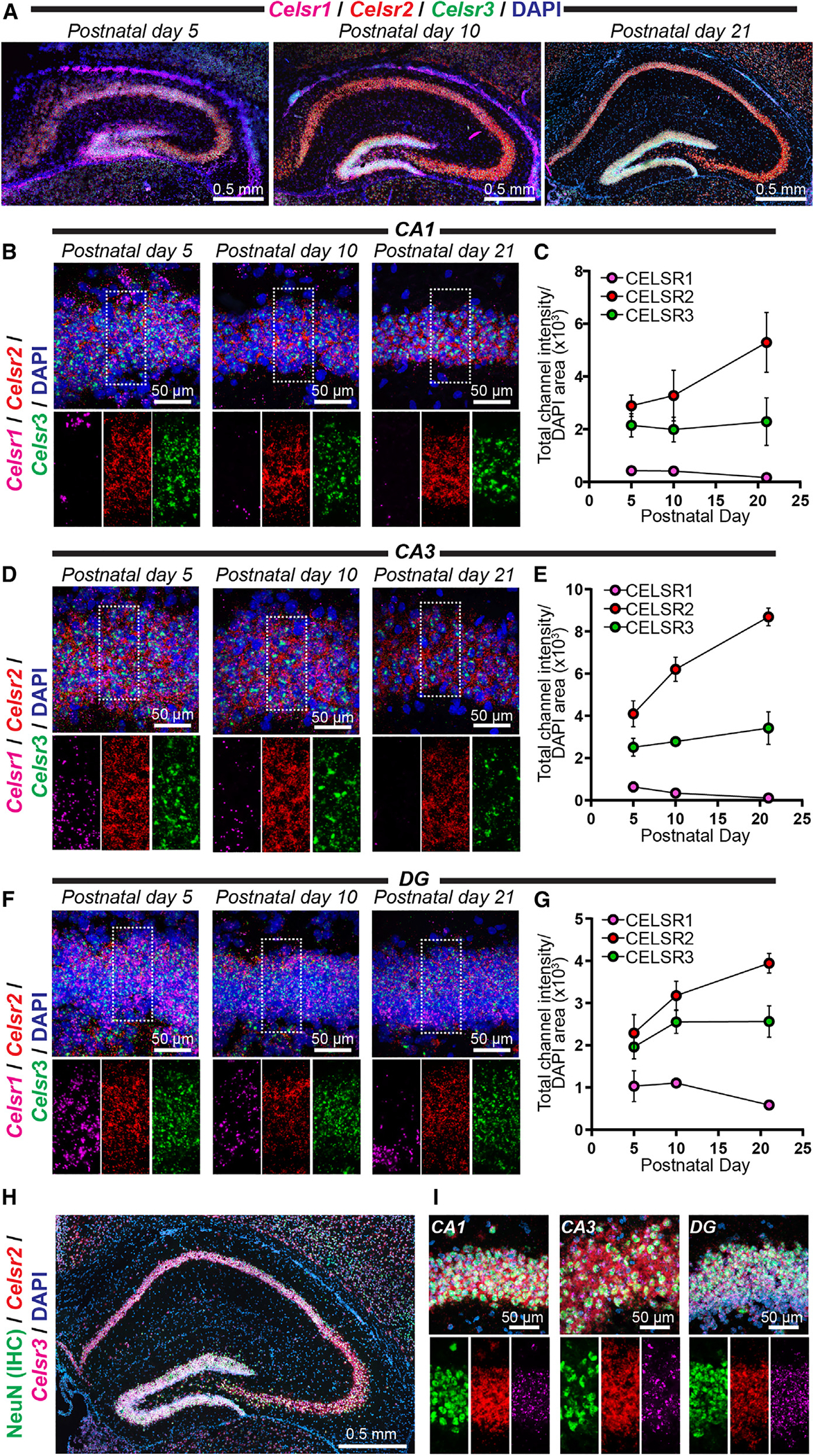
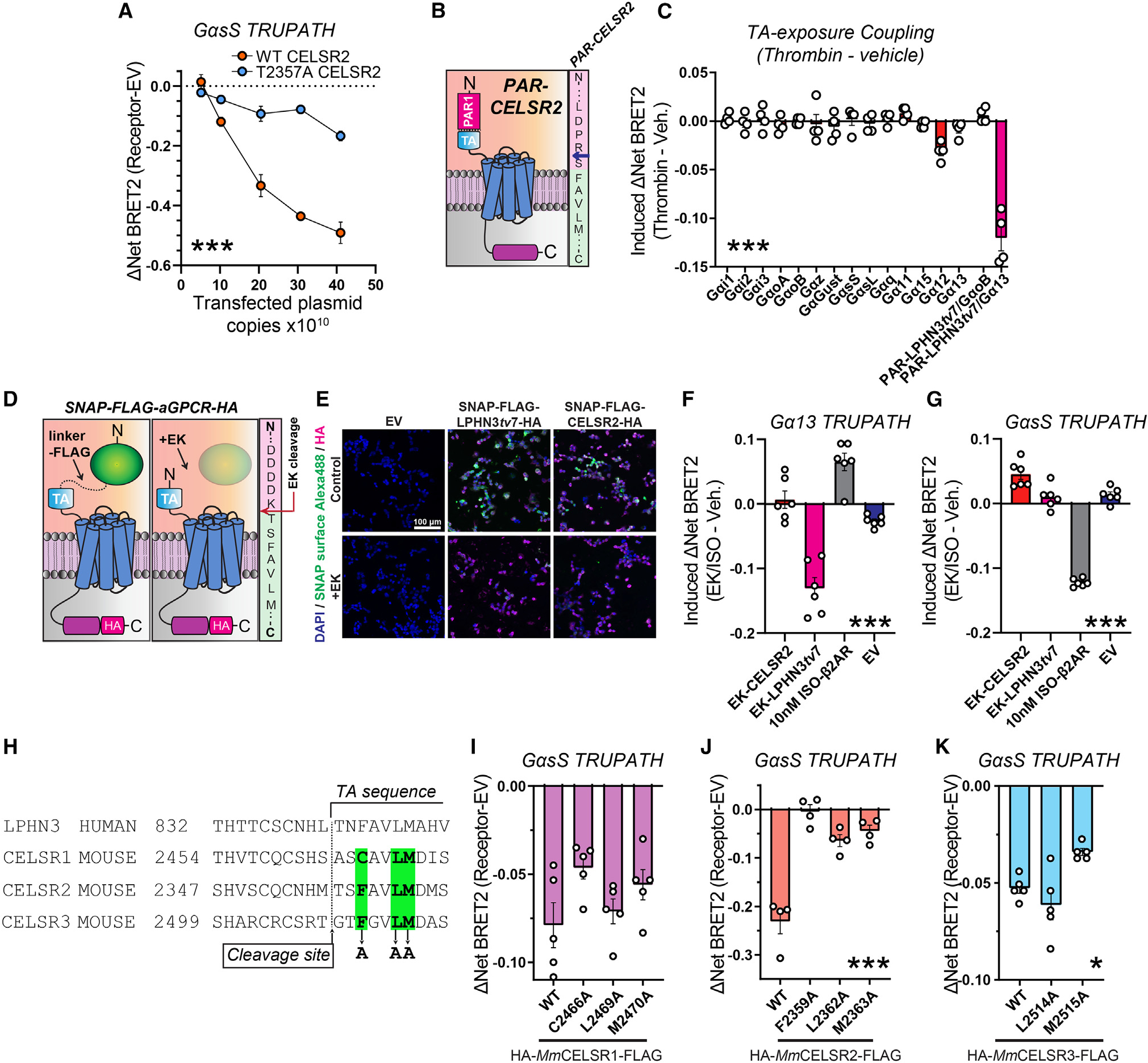
Adhesion G protein-coupled receptors (aGPCRs) are a large GPCR class that direct diverse fundamental biological processes. One prominent mechanism for aGPCR agonism involves autoproteolytic cleavage, which generates an activating, membrane-proximal tethered agonist (TA). How universal this mechanism is for all aGPCRs is unclear. Here, we investigate G protein induction principles of aGPCRs using mammalian latrophilin 3 (LPHN3) and cadherin EGF LAG-repeat 7-transmembrane receptors 1-3 (CELSR1-3), members of two aGPCR families conserved from invertebrates to vertebrates. LPHNs and CELSRs mediate fundamental aspects of brain development, yet CELSR signaling mechanisms are unknown. We find that CELSR1 and CELSR3 are cleavage deficient, while CELSR2 is efficiently cleaved. Despite differential autoproteolysis, CELSR1-3 all engage GαS, and CELSR1 or CELSR3 TA point mutants retain GαS coupling activity. CELSR2 autoproteolysis enhances GαS coupling, yet acute TA exposure alone is insufficient. These studies support that aGPCRs signal via multiple paradigms and provide insights into CELSR biological function.
Keywords: CP: Cell biology; adhesion GPCR; autoproteolysis; cell adhesion; signal transduction.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Update of
-
The adhesion GPCRs CELSR1-3 and LPHN3 engage G proteins via distinct activation mechanisms.bioRxiv [Preprint]. 2023 Apr 3:2023.04.02.535287. doi: 10.1101/2023.04.02.535287. bioRxiv. 2023. Update in: Cell Rep. 2023 Jun 27;42(6):112552. doi: 10.1016/j.celrep.2023.112552. PMID: 37066404 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
