

Exercise metabolism and adaptation in skeletal muscle
- PMID: 37225892
- PMCID: PMC10527431
- DOI: 10.1038/s41580-023-00606-x
Exercise metabolism and adaptation in skeletal muscle
Abstract
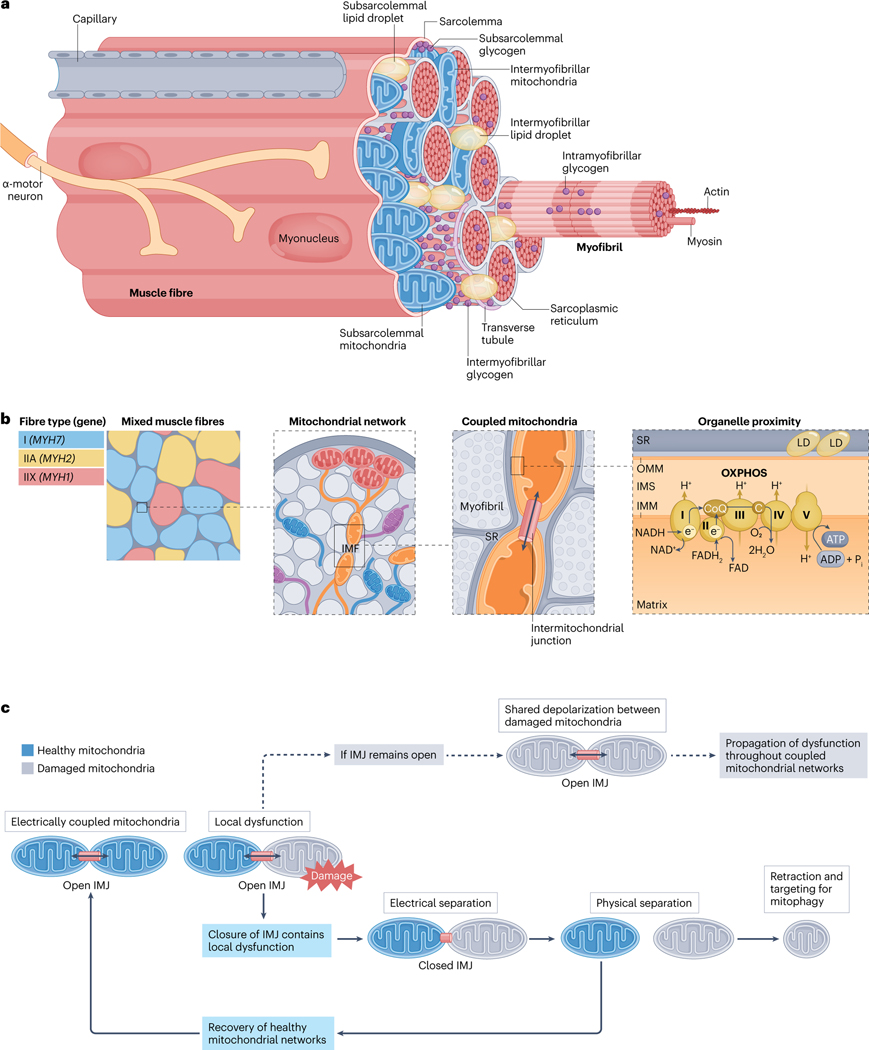
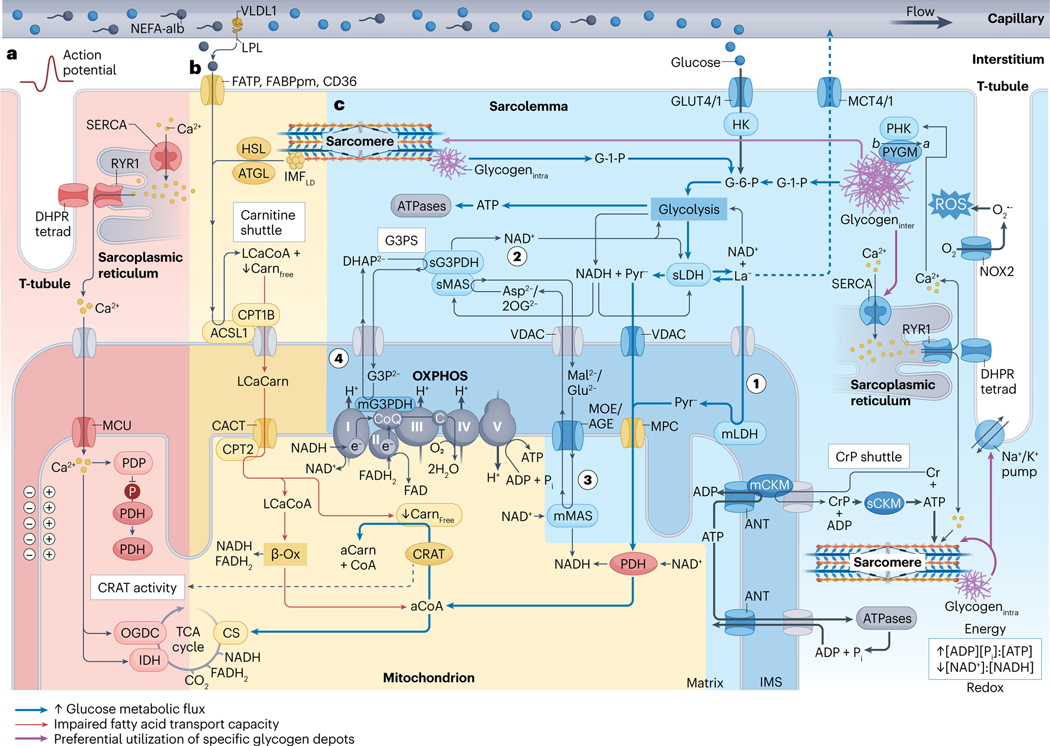
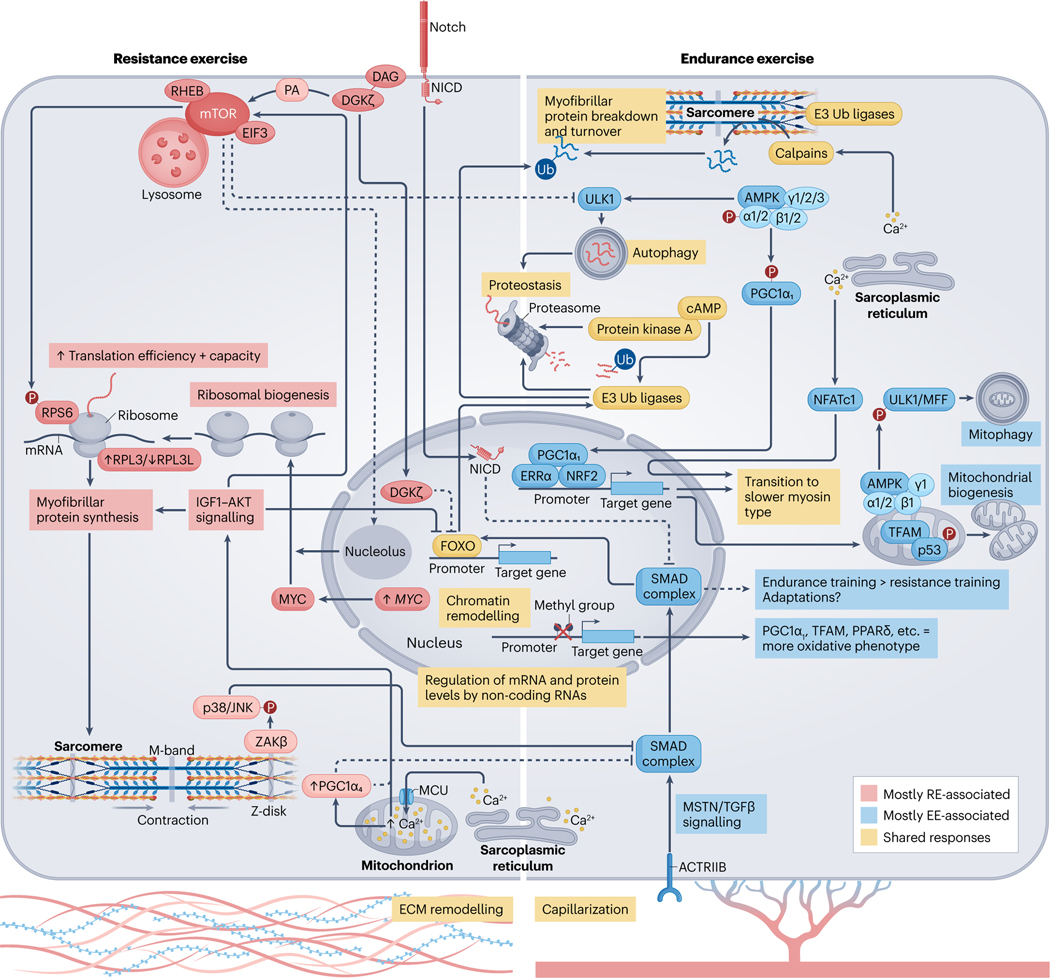
Viewing metabolism through the lens of exercise biology has proven an accessible and practical strategy to gain new insights into local and systemic metabolic regulation. Recent methodological developments have advanced understanding of the central role of skeletal muscle in many exercise-associated health benefits and have uncovered the molecular underpinnings driving adaptive responses to training regimens. In this Review, we provide a contemporary view of the metabolic flexibility and functional plasticity of skeletal muscle in response to exercise. First, we provide background on the macrostructure and ultrastructure of skeletal muscle fibres, highlighting the current understanding of sarcomeric networks and mitochondrial subpopulations. Next, we discuss acute exercise skeletal muscle metabolism and the signalling, transcriptional and epigenetic regulation of adaptations to exercise training. We address knowledge gaps throughout and propose future directions for the field. This Review contextualizes recent research of skeletal muscle exercise metabolism, framing further advances and translation into practice.
© 2023. Springer Nature Limited.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
-
- Bergouignan A. et al. Effect of contrasted levels of habitual physical activity on metabolic flexibility. J. Appl. Physiol 114, 371–379 (2013). - PubMed
-
- Zhang D. et al. Leisure-time physical activity and incident metabolic syndrome: a systematic review and dose-response meta-analysis of cohort studies. Metabolism 75, 36–44 (2017). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
