

Use of the dTAG system in vivo to degrade CDK2 and CDK5 in adult mice and explore potential safety liabilities
- PMID: 37228089
- PMCID: PMC10306401
- DOI: 10.1093/toxsci/kfad049
Use of the dTAG system in vivo to degrade CDK2 and CDK5 in adult mice and explore potential safety liabilities
Abstract
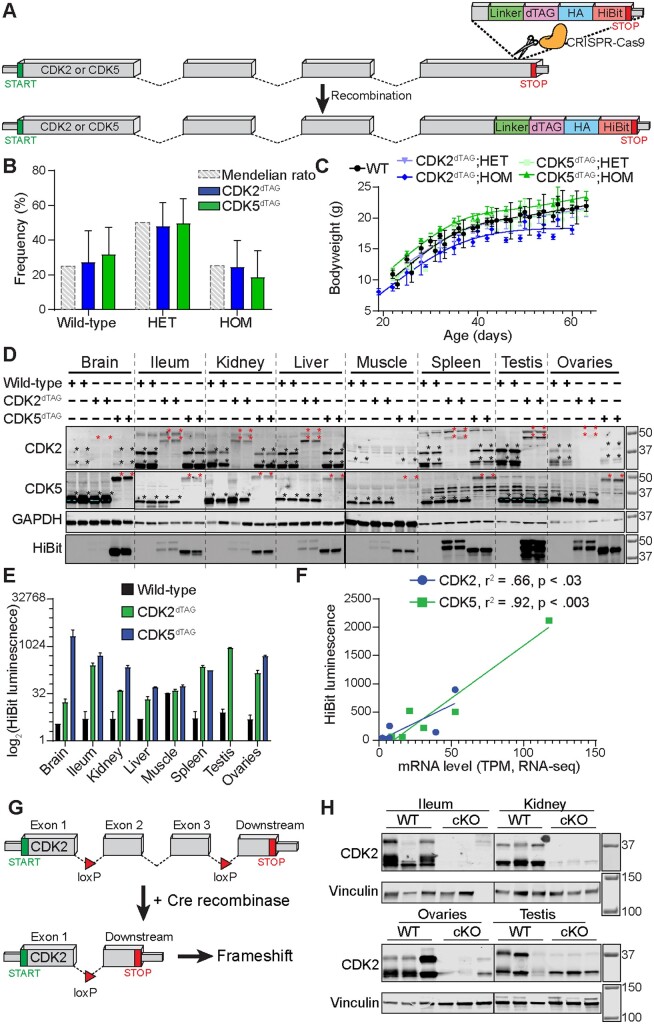
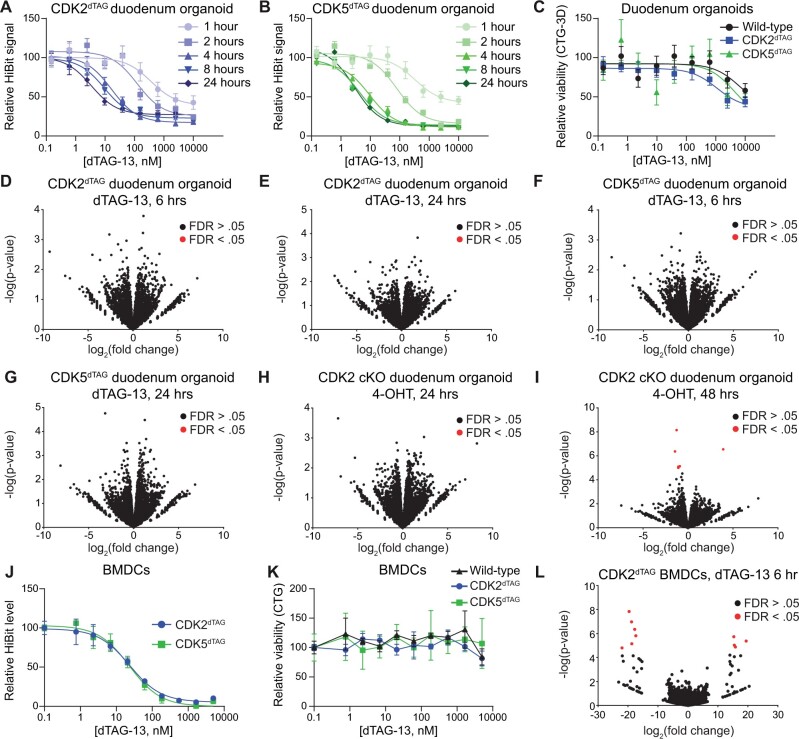
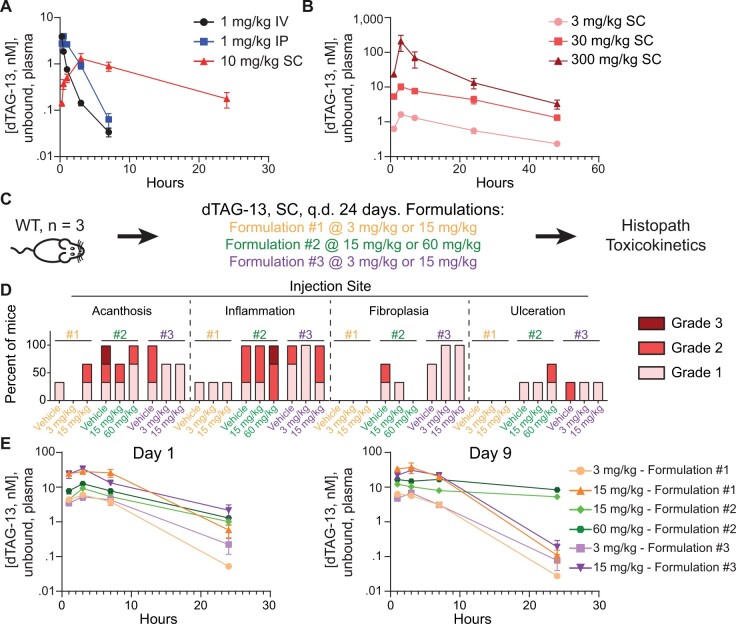
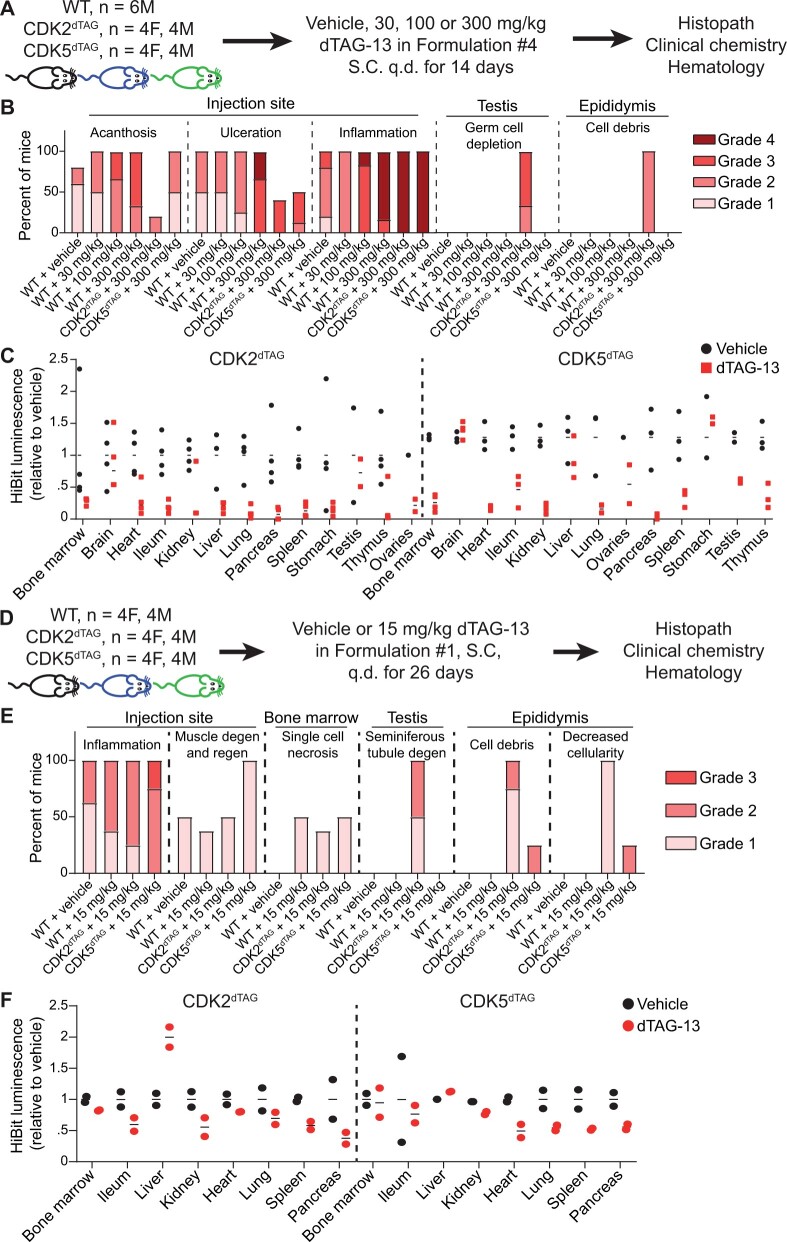
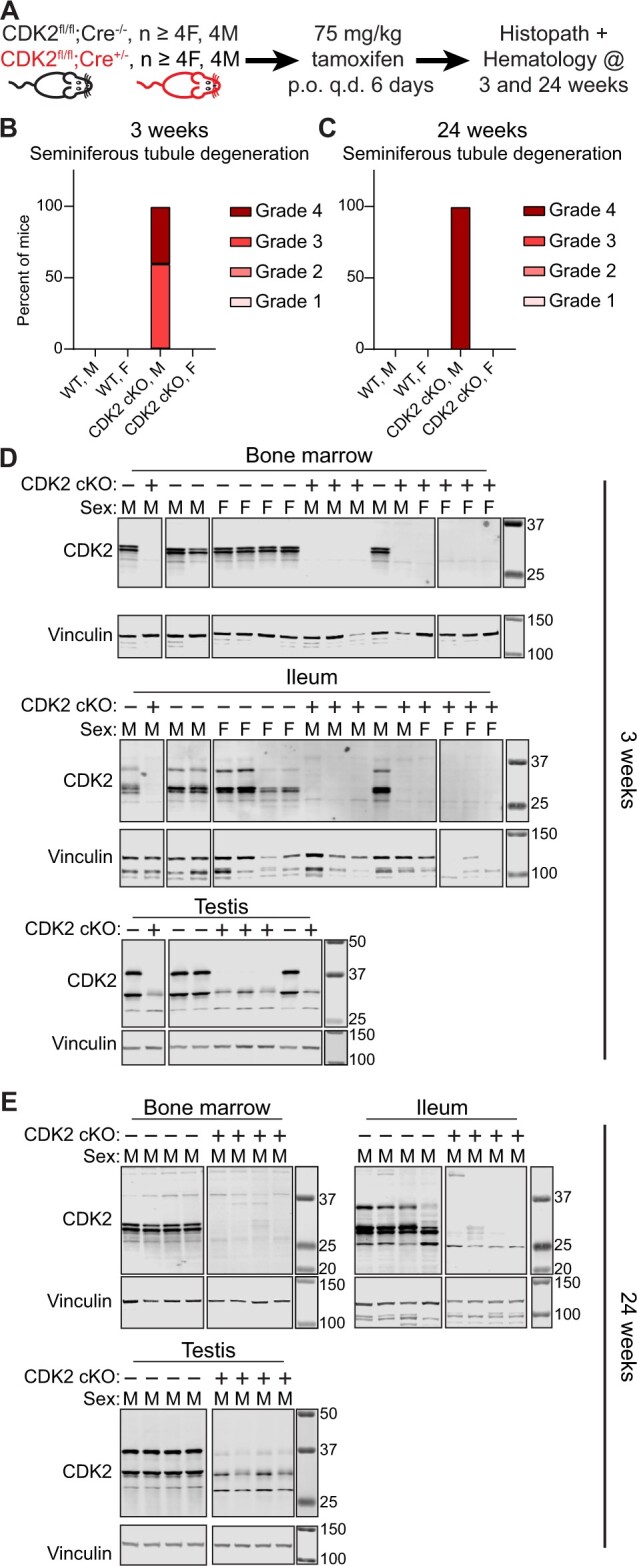
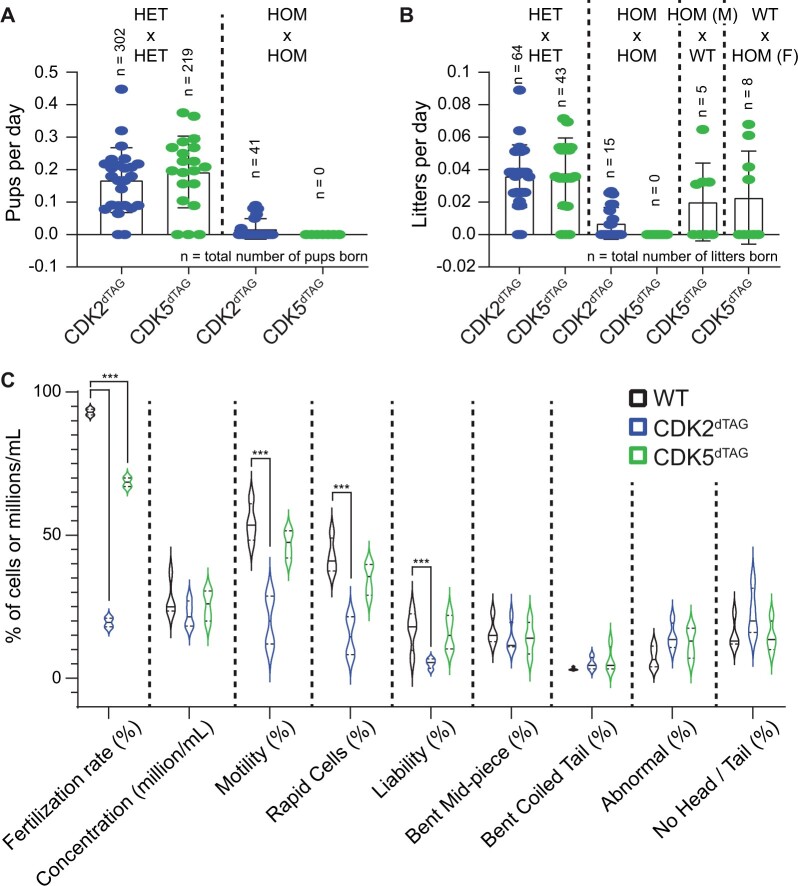
The degradation tag (dTAG) system for target protein degradation can remove proteins from biological systems without the drawbacks of some genetic methods, such as slow kinetics, lack of reversibility, low specificity, and the inability to titrate dosage. These drawbacks can make it difficult to compare toxicity resulting from genetic and pharmacological interventions, especially in vivo. Because the dTAG system has not been studied extensively in vivo, we explored the use of this system to study the physiological sequalae resulting from CDK2 or CDK5 degradation in adult mice. Mice with homozygous knock-in of the dTAG sequence onto CDK2 and CDK5 were born at Mendelian ratios despite decreased CDK2 or CDK5 protein levels in comparison with wild-type mice. In bone marrow cells and duodenum organoids derived from these mice, treatment with the dTAG degrader dTAG-13 resulted in rapid and robust protein degradation but caused no appreciable change in viability or the transcriptome. Repeated delivery of dTAG-13 in vivo for toxicity studies proved challenging; we explored multiple formulations in an effort to maximize degradation while minimizing formulation-related toxicity. Degradation of CDK2 or CDK5 in all organs except the brain, where dTAG-13 likely did not cross the blood brain barrier, only caused microscopic changes in the testis of CDK2dTAG mice. These findings were corroborated with conditional CDK2 knockout in adult mice. Our results suggest that the dTAG system can provide robust protein degradation in vivo and that loss of CDK2 or CDK5 in adult mice causes no previously unknown phenotypes.
Keywords: CDK2; CDK5; PROTAC; degradation; mice; toxicity.
© The Author(s) 2023. Published by Oxford University Press on behalf of the Society of Toxicology.
Figures
References
-
- Aulbach A., Vitsky A., Arndt T., Ramaiah L., Logan M., Siska W., Cregar L., Tripathi N., Adedeji A., Provencher A., et al. (2019). Interpretative considerations for clinical pathology findings in nonclinical toxicology studies. Vet. Clin. Pathol. 48, 383–388. - PubMed
-
- Berthet C., Aleem E., Coppola V., Tessarollo L., Kaldis P. (2003). CDK2 knockout mice are viable. Curr. Biol. 13, 1775–1785. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
