

Amplification is the primary mode of gene-by-sex interaction in complex human traits
- PMID: 37228747
- PMCID: PMC10203050
- DOI: 10.1016/j.xgen.2023.100297
Amplification is the primary mode of gene-by-sex interaction in complex human traits
Abstract
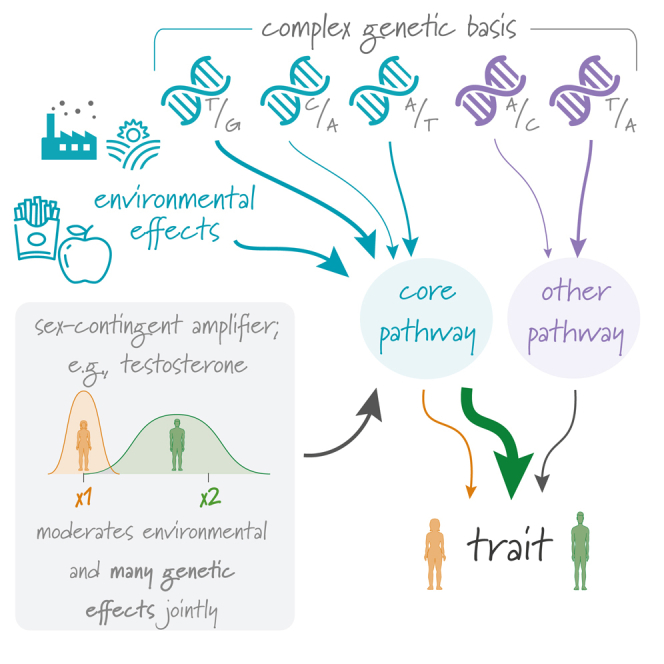
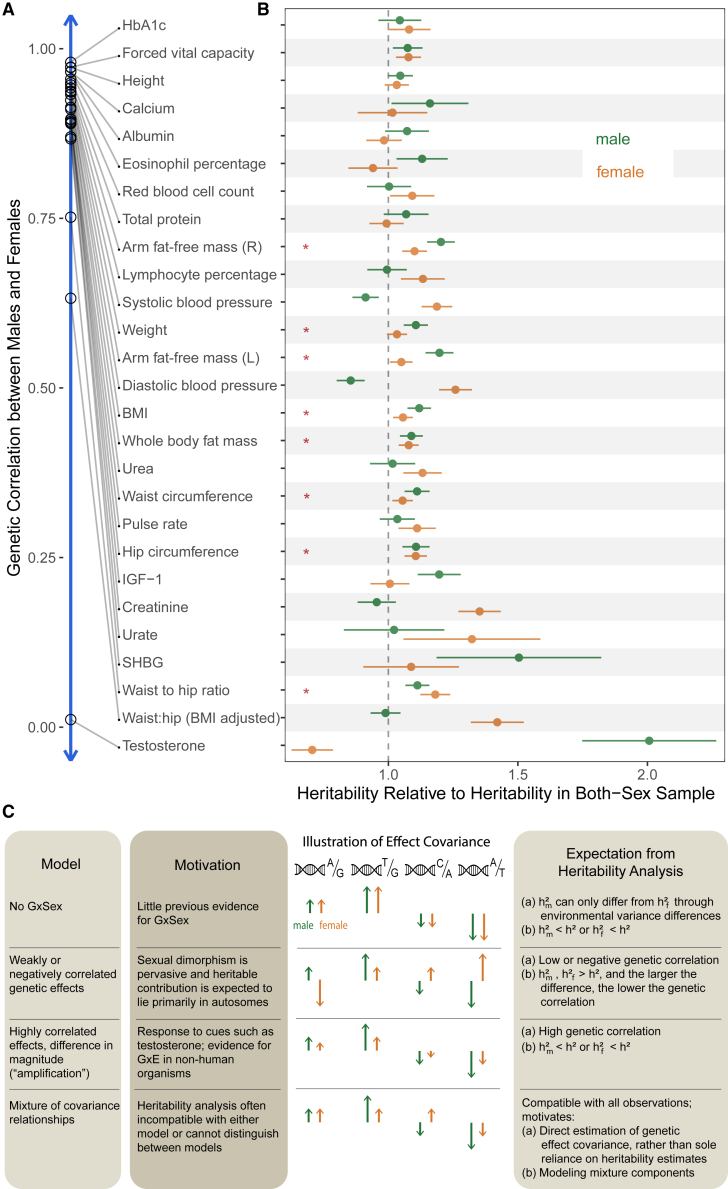
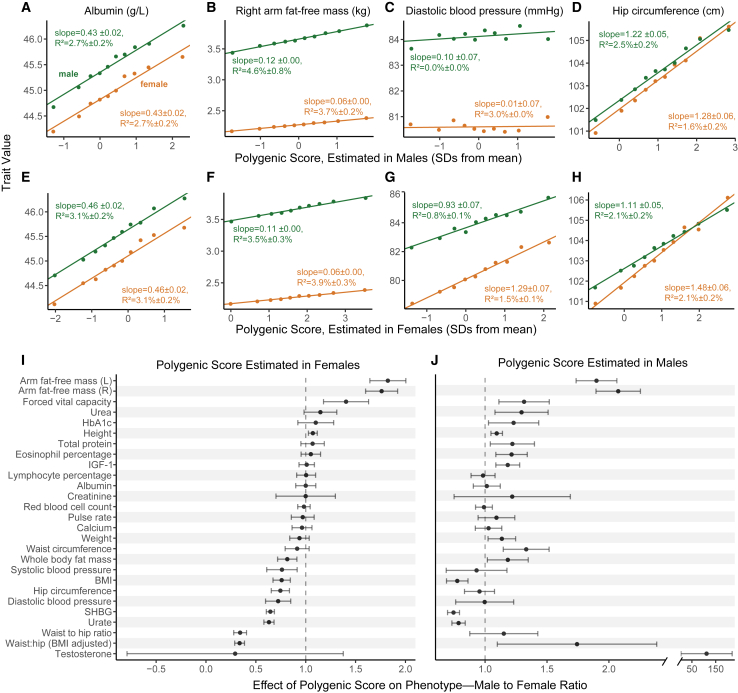
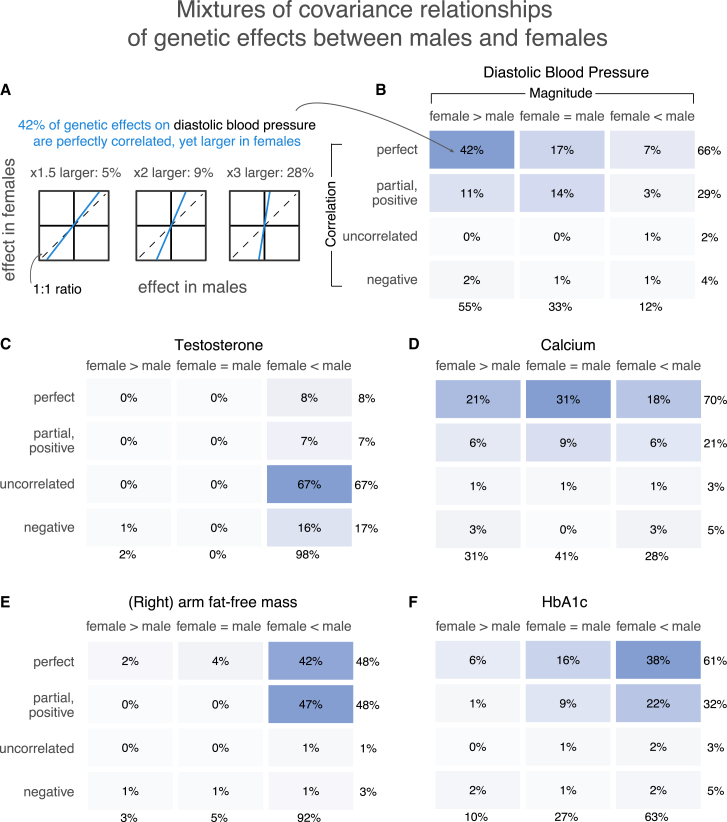
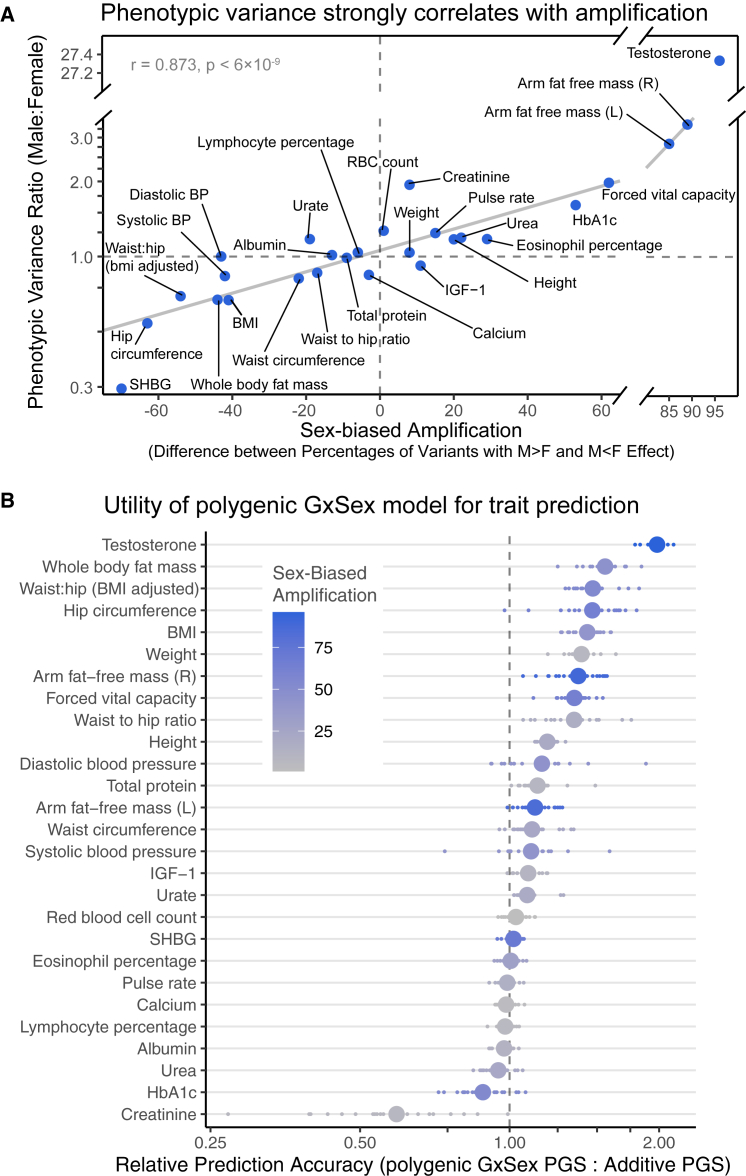
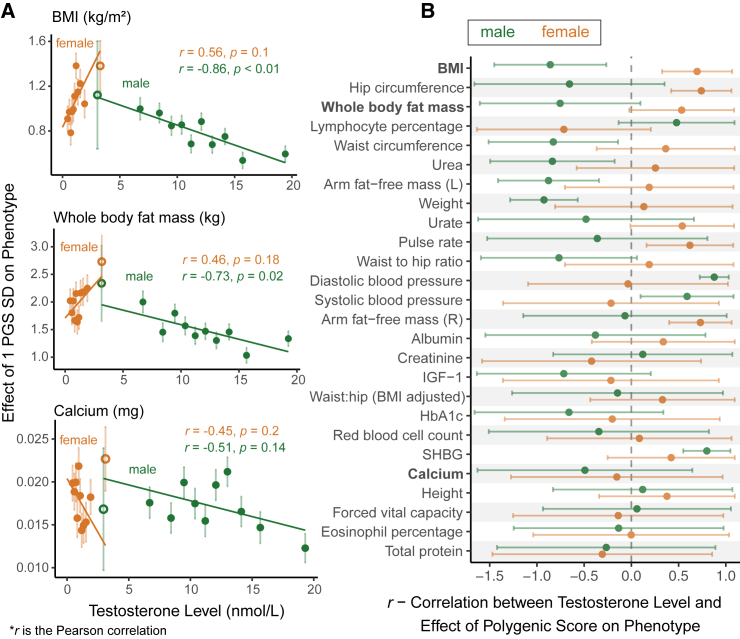
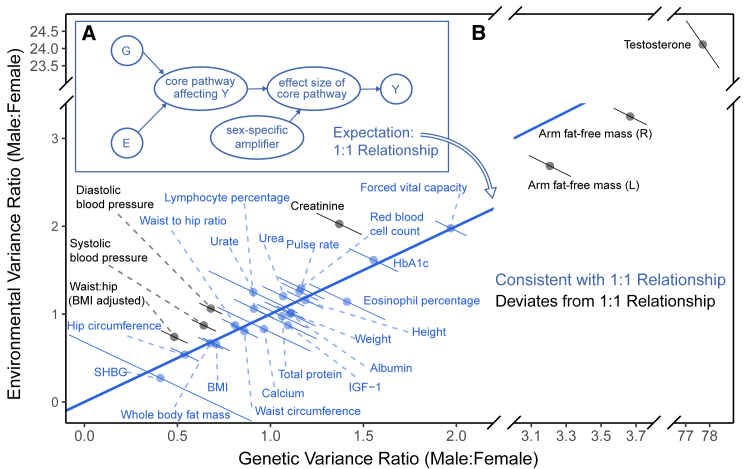
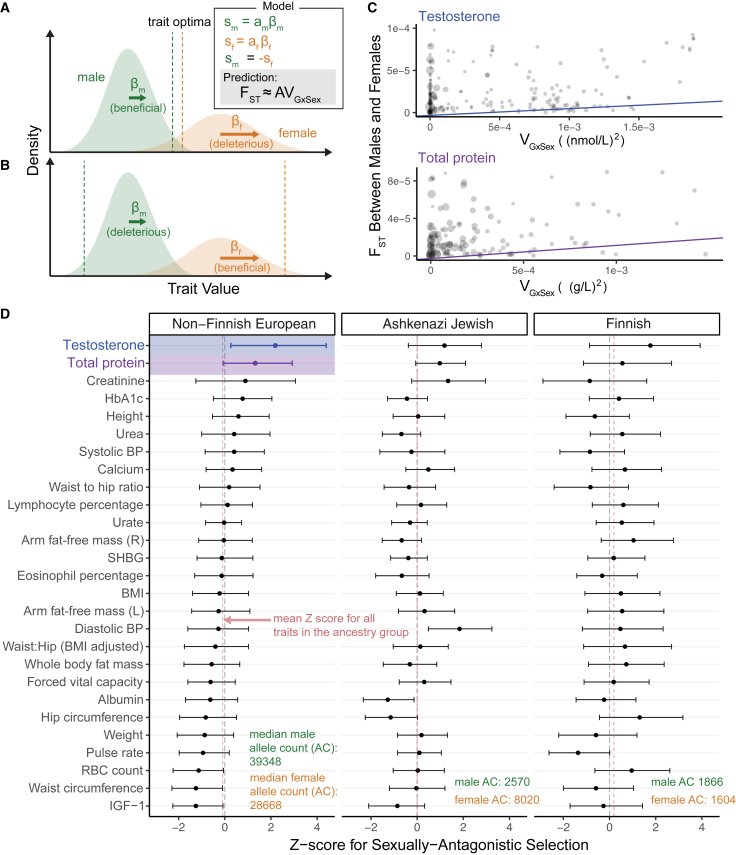
Sex differences in complex traits are suspected to be in part due to widespread gene-by-sex interactions (GxSex), but empirical evidence has been elusive. Here, we infer the mixture of ways in which polygenic effects on physiological traits covary between males and females. We find that GxSex is pervasive but acts primarily through systematic sex differences in the magnitude of many genetic effects ("amplification") rather than in the identity of causal variants. Amplification patterns account for sex differences in trait variance. In some cases, testosterone may mediate amplification. Finally, we develop a population-genetic test linking GxSex to contemporary natural selection and find evidence of sexually antagonistic selection on variants affecting testosterone levels. Our results suggest that amplification of polygenic effects is a common mode of GxSex that may contribute to sex differences and fuel their evolution.
© 2023 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Arnqvist G., Rowe L. Princeton University Press; 2005. Sexual Conflict. - DOI
-
- Bayer E.A., Stecky R.C., Neal L., Katsamba P.S., Ahlsen G., Balaji V., Hoppe T., Shapiro L., Oren-Suissa M., Hobert O. Ubiquitin-dependent regulation of a conserved DMRT protein controls sexually dimorphic synaptic connectivity and behavior. Elife. 2020;9 doi: 10.7554/eLife.59614. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
