

β-Sitosterol as a Promising Anticancer Agent for Chemoprevention and Chemotherapy: Mechanisms of Action and Future Prospects
- PMID: 37247842
- PMCID: PMC10509430
- DOI: 10.1016/j.advnut.2023.05.013
β-Sitosterol as a Promising Anticancer Agent for Chemoprevention and Chemotherapy: Mechanisms of Action and Future Prospects
Abstract
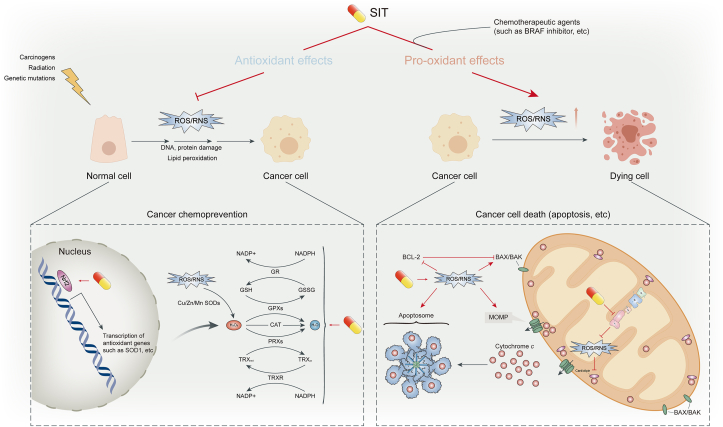
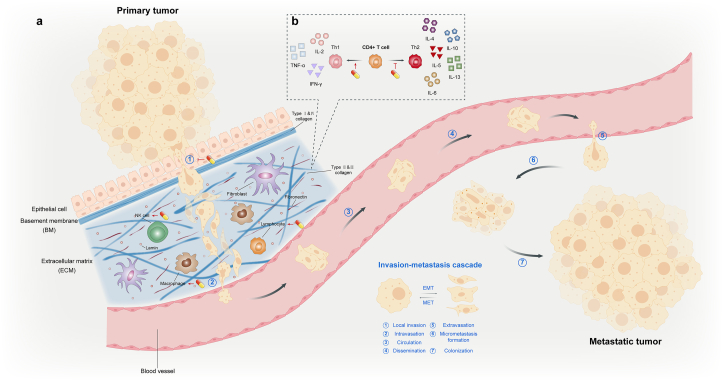
Cancer is one of the primary causes of death worldwide, and its incidence continues to increase yearly. Despite significant advances in research, the search for effective and nontoxic preventive and therapeutic agents remains greatly important. Cancer is a multimodal disease, where various mechanisms play significant roles in its occurrence and progression. This highlights the need for multitargeted approaches that are not only safe and inexpensive but also provide effective alternatives for current therapeutic regimens. β-Sitosterol (SIT), the most abundant phytosterol found in various plant foods, represents such an option. Preclinical evidence over the past few decades has overwhelmingly shown that SIT exhibits multiple anticancer activities against varied cancers, such as liver, cervical, colon, stomach, breast, lung, pancreatic, and prostate cancers, in addition to leukemia, multiple myeloma, melanoma, and fibrosarcoma. In this article, we present the latest advances and perspectives on SIT-systematically summarizing its antitumor mechanisms of action into 7 main sections and combining current challenges and prospects-for its use as a promising agent for cancer prevention and treatment. In particular, SIT plays a role in cancer prevention and treatment mainly by enhancing apoptosis, inducing cell cycle arrest, bidirectionally regulating oxidative stress, improving metabolic reprogramming, inhibiting invasion and metastasis, modulating immunity and inflammation, and combating drug resistance. Although SIT holds such great promise, the poor aqueous solubility and bioavailability coupled with low targeting efficacy limit its therapeutic efficacy and clinical application. Further research on novel drug delivery systems may improve these deficiencies. Overall, through complex and pleiotropic mechanisms, SIT has good potential for tumor chemoprevention and chemotherapy. However, no clinical trials have yet proven this potential. This review provides theoretical basis and rationality for the further design and conduct of clinical trials to confirm the anticancer activity of SIT.
Keywords: cancer; chemoprevention; chemotherapy; phytochemical; phytosterol; β-sitosterol.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Figures





References
-
- Wu C., Li M., Meng H., Liu Y., Niu W., Zhou Y., et al. Analysis of status and countermeasures of cancer incidence and mortality in China. Sci. China Life. Sci. 2019;62:640–647. - PubMed
-
- Sung H., Ferlay J., Siegel R.L., Laversanne M., Soerjomataram I., Jemal A., et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. C.A. Cancer J. Clin. 2021;71:209–249. - PubMed
-
- Buyel J.F. Plants as sources of natural and recombinant anti-cancer agents. Biotechnol. Adv. 2018;36:506–520. - PubMed
-
- Nasir A., Bullo M.M.H., Ahmed Z., Imtiaz A., Yaqoob E., Jadoon M., et al. Nutrigenomics: epigenetics and cancer prevention: a comprehensive review. Crit. Rev. Food Sci. Nutr. 2020;60:1375–1387. - PubMed
-
- Moreau R.A., Whitaker B.D., Hicks K.B. Phytosterols, phytostanols, and their conjugates in foods: structural diversity, quantitative analysis, and health-promoting uses. Prog. Lipid Res. 2002;41:457–500. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
