

Postsynaptic synucleins mediate endocannabinoid signaling
- PMID: 37248337
- PMCID: PMC10244176
- DOI: 10.1038/s41593-023-01345-0
Postsynaptic synucleins mediate endocannabinoid signaling
Abstract
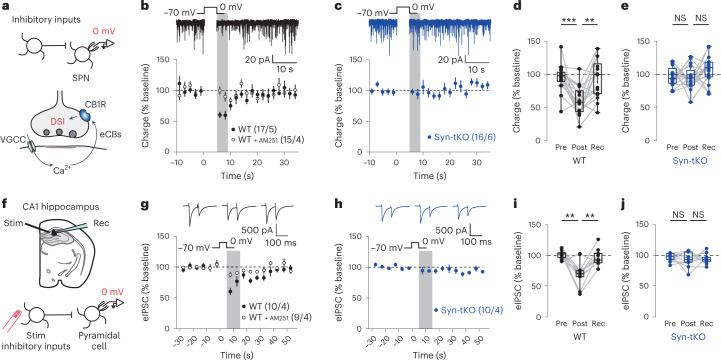
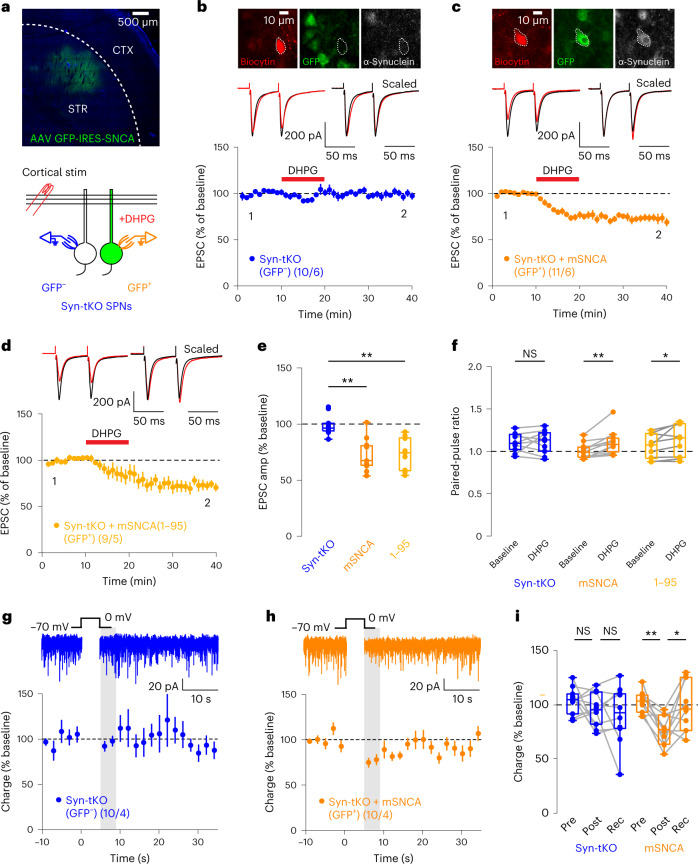
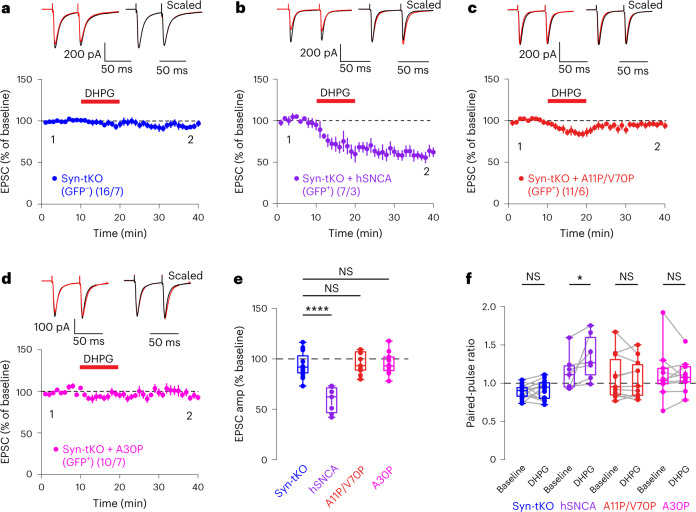
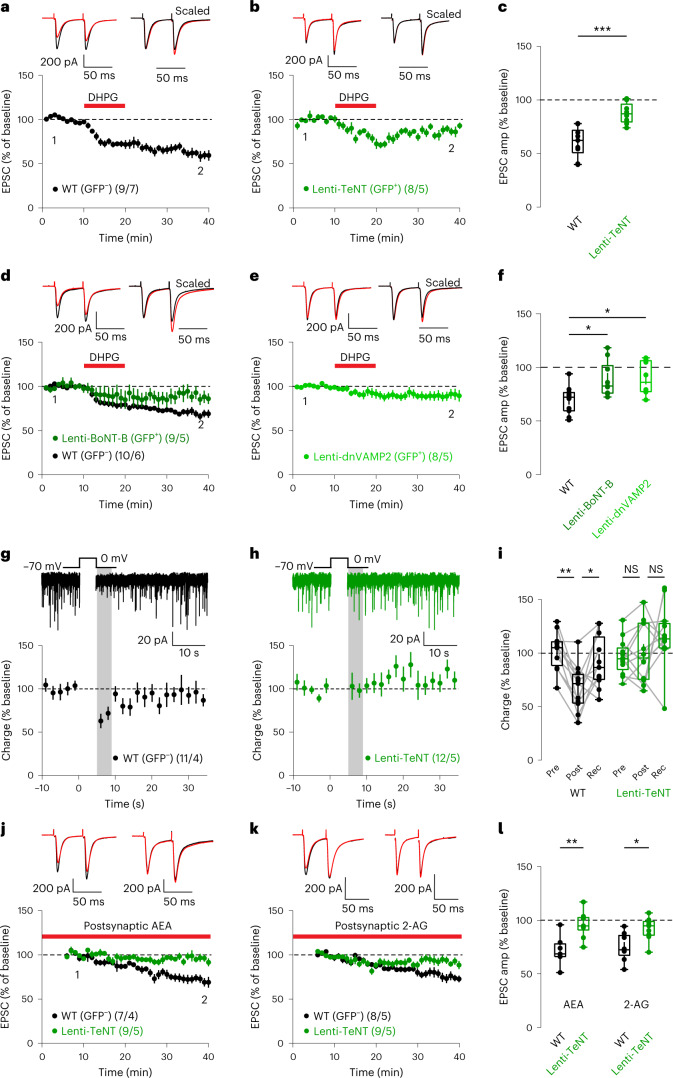
Endocannabinoids are among the most powerful modulators of synaptic transmission throughout the nervous system, and yet little is understood about the release of endocannabinoids from postsynaptic compartments. Here we report an unexpected finding that endocannabinoid release requires synucleins, key contributors to Parkinson's disease. We show that endocannabinoids are released postsynaptically by a synuclein-dependent and SNARE-dependent mechanism. Specifically, we found that synuclein deletion blocks endocannabinoid-dependent synaptic plasticity; this block is reversed by postsynaptic expression of wild-type but not of mutant α-synuclein. Whole-cell recordings and direct optical monitoring of endocannabinoid signaling suggest that the synuclein deletion specifically blocks endocannabinoid release. Given the presynaptic role of synucleins in regulating vesicle lifecycle, we hypothesize that endocannabinoids are released via a membrane interaction mechanism. Consistent with this hypothesis, postsynaptic expression of tetanus toxin light chain, which cleaves synaptobrevin SNAREs, also blocks endocannabinoid-dependent signaling. The unexpected finding that endocannabinoids are released via a synuclein-dependent mechanism is consistent with a general function of synucleins in membrane trafficking and adds a piece to the longstanding puzzle of how neurons release endocannabinoids to induce synaptic plasticity.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures











References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
