

Aberrant activation of Notch1 signaling in the mouse uterine epithelium promotes hyper-proliferation by increasing estrogen sensitivity
- PMID: 37249327
- PMCID: PMC10263383
- DOI: 10.1096/fj.202201868RR
Aberrant activation of Notch1 signaling in the mouse uterine epithelium promotes hyper-proliferation by increasing estrogen sensitivity
Abstract
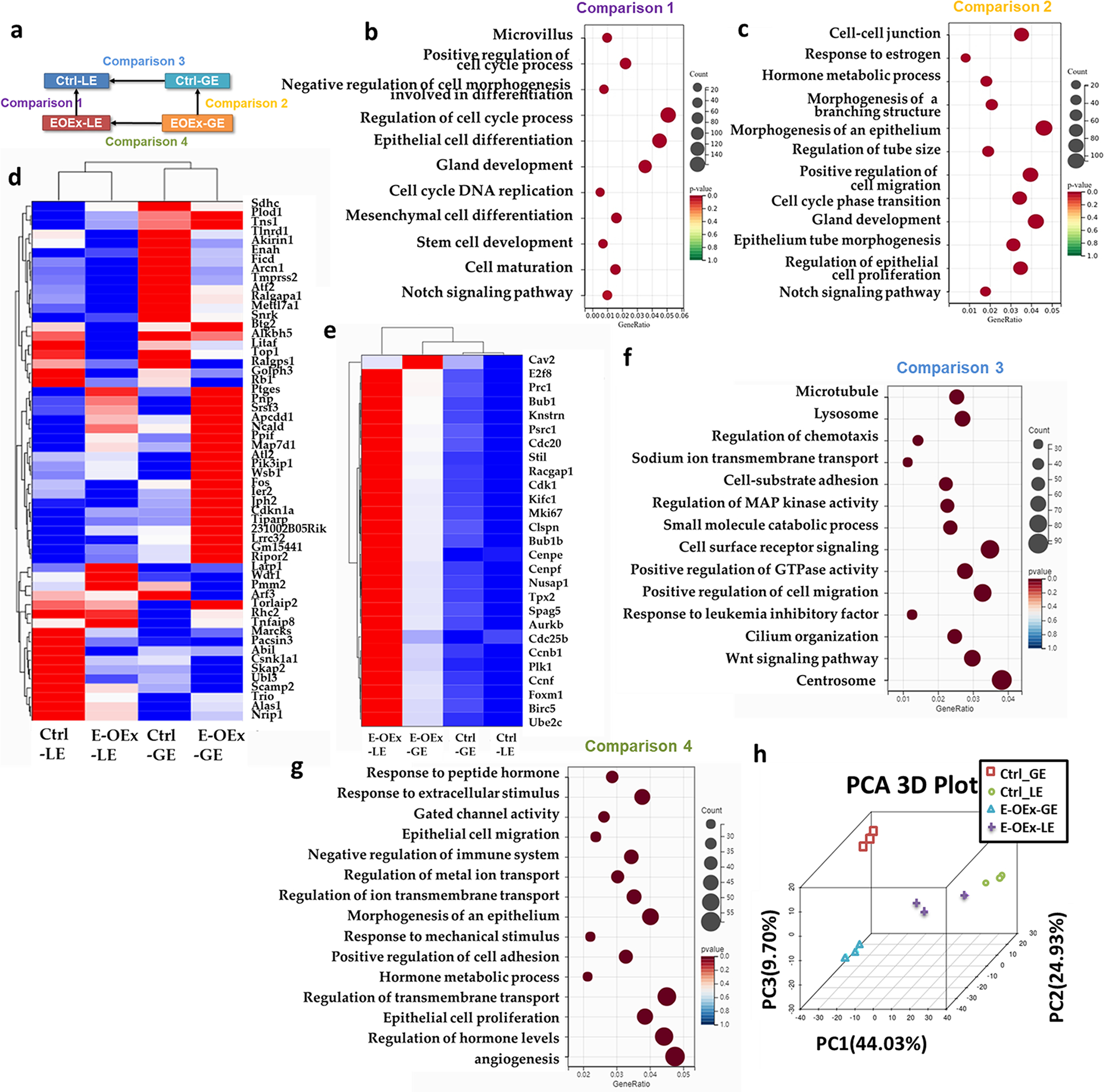
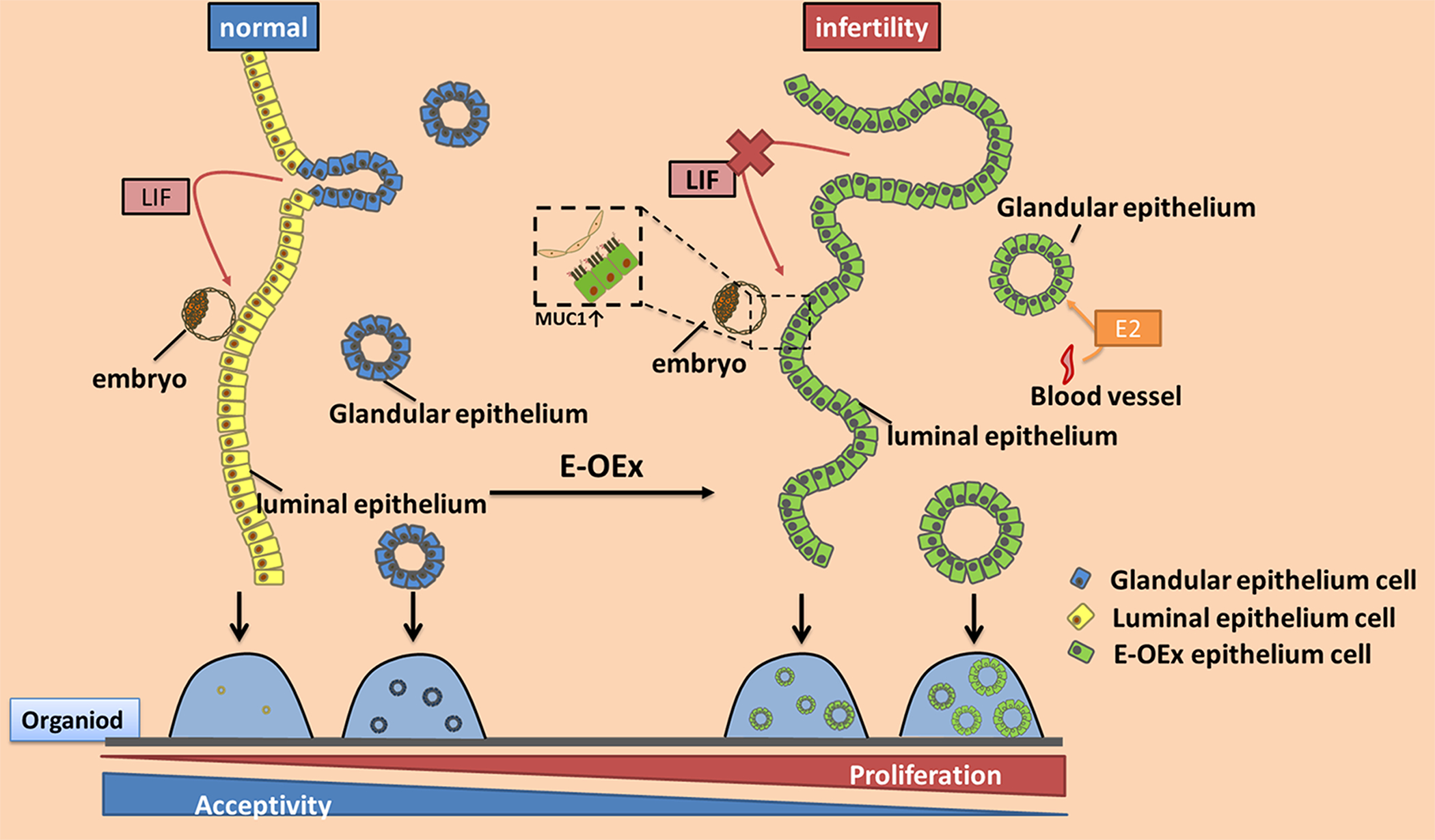
In mammals, the endometrium undergoes dynamic changes in response to estrogen and progesterone to prepare for blastocyst implantation. Two distinct types of endometrial epithelial cells, the luminal (LE) and glandular (GE) epithelial cells play different functional roles during this physiological process. Previously, we have reported that Notch signaling plays multiple roles in embryo implantation, decidualization, and postpartum repair. Here, using the uterine epithelial-specific Ltf-iCre, we showed that Notch1 signaling over-activation in the endometrial epithelium caused dysfunction of the epithelium during the estrous cycle, resulting in hyper-proliferation. During pregnancy, it further led to dysregulation of estrogen and progesterone signaling, resulting in infertility in these animals. Using 3D organoids, we showed that over-activation of Notch1 signaling increased the proliferative potential of both LE and GE cells and reduced the difference in transcription profiles between them, suggesting disrupted differentiation of the uterine epithelium. In addition, we demonstrated that both canonical and non-canonical Notch signaling contributed to the hyper-proliferation of GE cells, but only the non-canonical pathway was involved with estrogen sensitivity in the GE cells. These findings provided insights into the effects of Notch1 signaling on the proliferation, differentiation, and function of the uterine epithelium. This study demonstrated the important roles of Notch1 signaling in regulating hormone response and differentiation of endometrial epithelial cells and provides an opportunity for future studies in estrogen-dependent diseases, such as endometriosis.
Keywords: Notch1 signaling; estrogen sensitivity; glandular epithelium; hyper-proliferation; luminal epithelium.
© 2023 The Authors. The FASEB Journal published by Wiley Periodicals LLC on behalf of Federation of American Societies for Experimental Biology.
Conflict of interest statement
Conflict of Interest
The authors declare no competing financial interests.
Figures




References
-
- Wood GA, Fata JE, Watson KLM and Khokha R Circulating hormones and estrous stage predict cellular and stromal remodeling in murine uterus. Reproduction. 2007; 133: 1035–1044. - PubMed
-
- Habiba M, Heyn R, Bianchi P, Brosens I and Benagiano G The development of the human uterus: morphogenesis to menarche. Hum Reprod Update. 2021; 27: 1–26. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
