

Dihydroxy-Metabolites of Dihomo-γ-linolenic Acid Drive Ferroptosis-Mediated Neurodegeneration
- PMID: 37252355
- PMCID: PMC10214511
- DOI: 10.1021/acscentsci.3c00052
Dihydroxy-Metabolites of Dihomo-γ-linolenic Acid Drive Ferroptosis-Mediated Neurodegeneration
Abstract
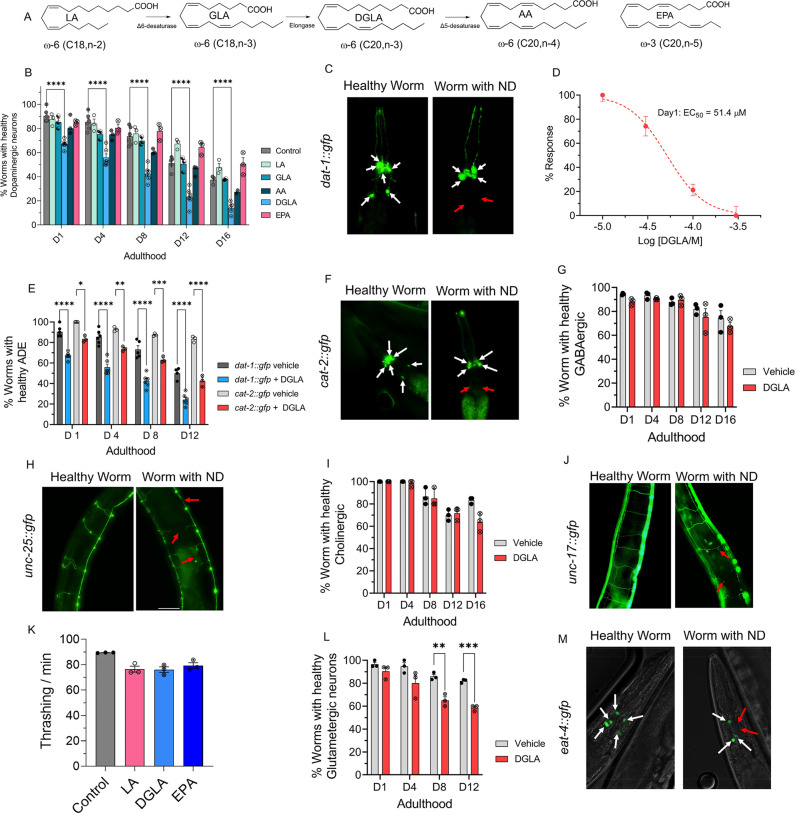
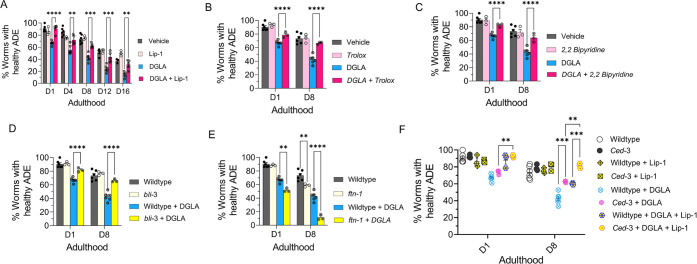
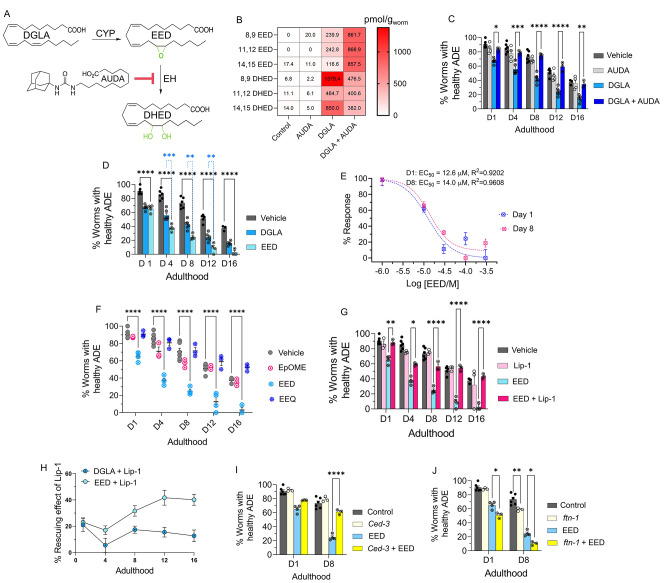
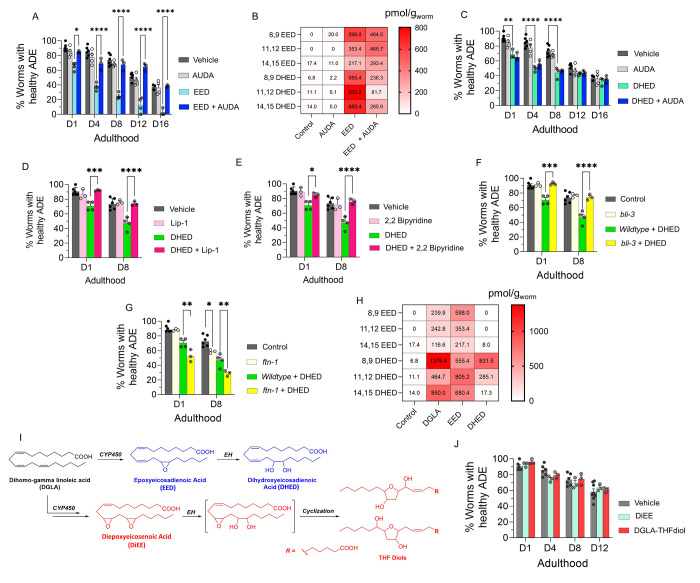
Even after decades of research, the mechanism of neurodegeneration remains understudied, hindering the discovery of effective treatments for neurodegenerative diseases. Recent reports suggest that ferroptosis could be a novel therapeutic target for neurodegenerative diseases. While polyunsaturated fatty acid (PUFA) plays an important role in neurodegeneration and ferroptosis, how PUFAs may trigger these processes remains largely unknown. PUFA metabolites from cytochrome P450 and epoxide hydrolase metabolic pathways may modulate neurodegeneration. Here, we test the hypothesis that specific PUFAs regulate neurodegeneration through the action of their downstream metabolites by affecting ferroptosis. We find that the PUFA dihomo-γ-linolenic acid (DGLA) specifically induces ferroptosis-mediated neurodegeneration in dopaminergic neurons. Using synthetic chemical probes, targeted metabolomics, and genetic mutants, we show that DGLA triggers neurodegeneration upon conversion to dihydroxyeicosadienoic acid through the action of CYP-EH (CYP, cytochrome P450; EH, epoxide hydrolase), representing a new class of lipid metabolites that induce neurodegeneration via ferroptosis.
© 2023 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Update of
-
Dihydroxy-Metabolites of Dihomo-gamma-linolenic Acid Drive Ferroptosis-Mediated Neurodegeneration.bioRxiv [Preprint]. 2023 Jan 10:2023.01.05.522933. doi: 10.1101/2023.01.05.522933. bioRxiv. 2023. Update in: ACS Cent Sci. 2023 Mar 16;9(5):870-882. doi: 10.1021/acscentsci.3c00052. PMID: 36711920 Free PMC article. Updated. Preprint.
References
-
- United Nations Department of Economic and Social Affairs . References. In World Population Ageing 2019; UN, 2020; pp 37–38.10.18356/7f55a58e-en. - DOI
-
- Dixon S. J.; Lemberg K. M.; Lamprecht M. R.; Skouta R.; Zaitsev E. M.; Gleason C. E.; Patel D. N.; Bauer A. J.; Cantley A. M.; Yang W. S.; Morrison B.; Stockwell B. R. Ferroptosis: An Iron-Dependent Form of Nonapoptotic Cell Death. Cell 2012, 149 (5), 1060–1072. 10.1016/j.cell.2012.03.042. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
