

Tuft cells mediate commensal remodeling of the small intestinal antimicrobial landscape
- PMID: 37253002
- PMCID: PMC10266004
- DOI: 10.1073/pnas.2216908120
Tuft cells mediate commensal remodeling of the small intestinal antimicrobial landscape
Abstract
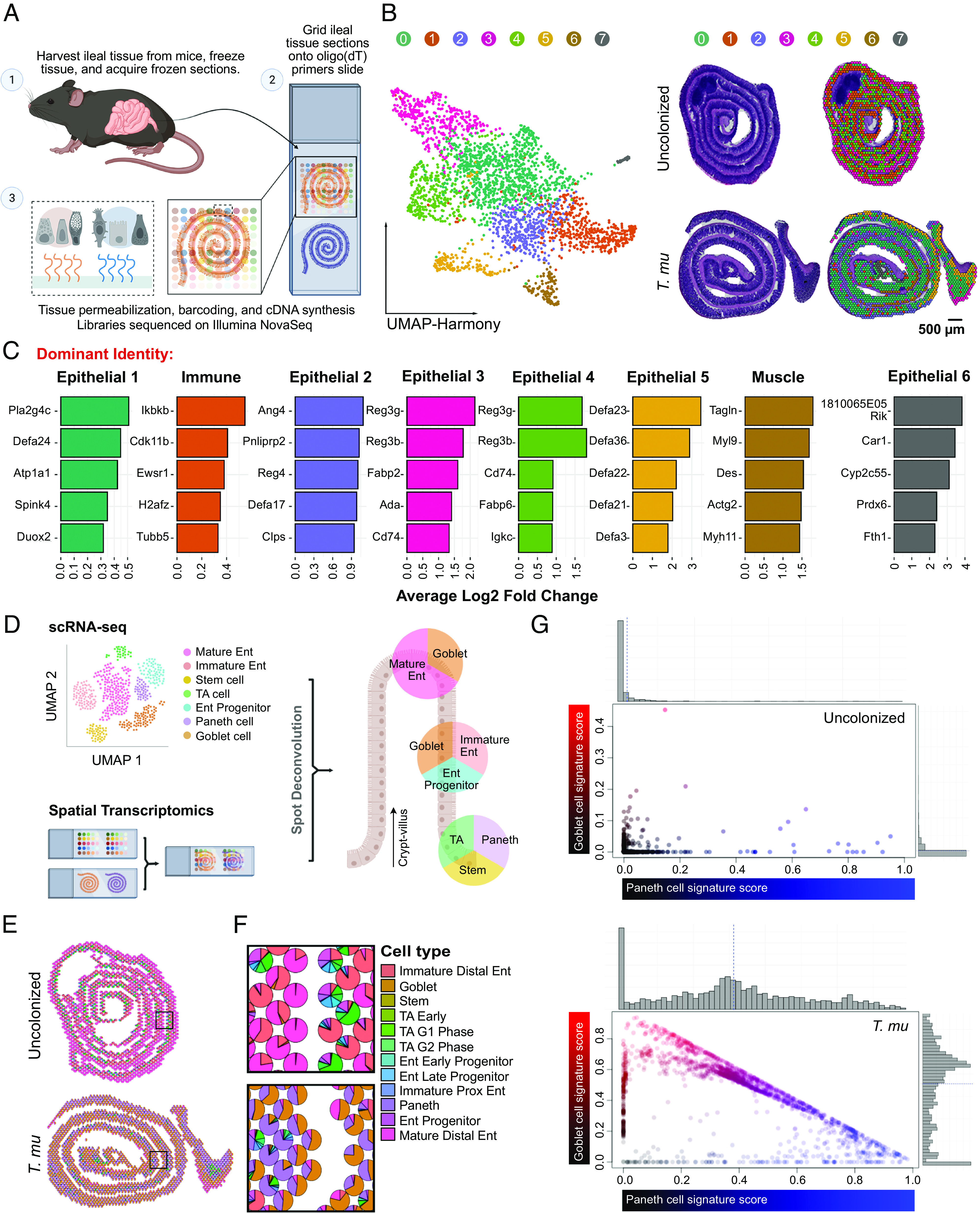
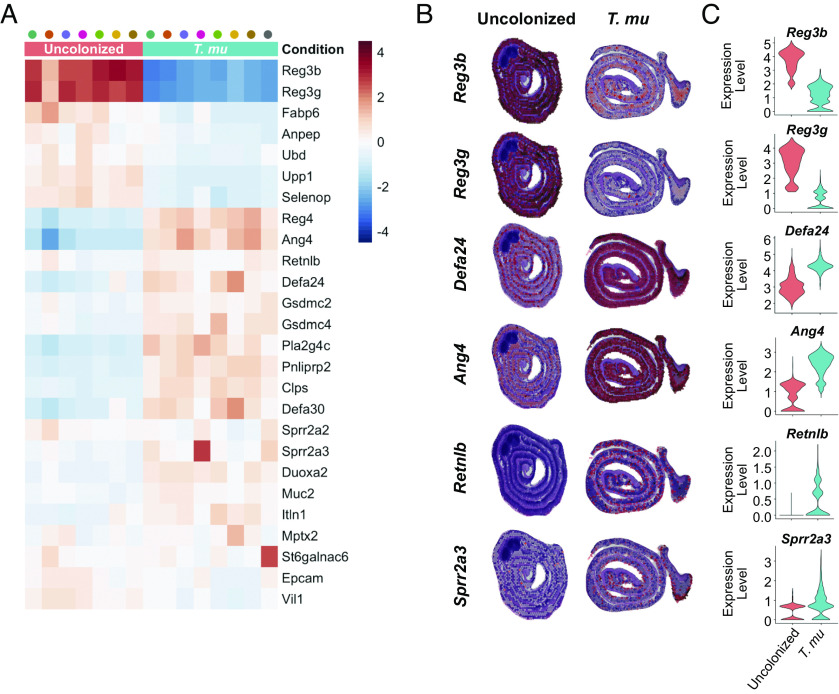
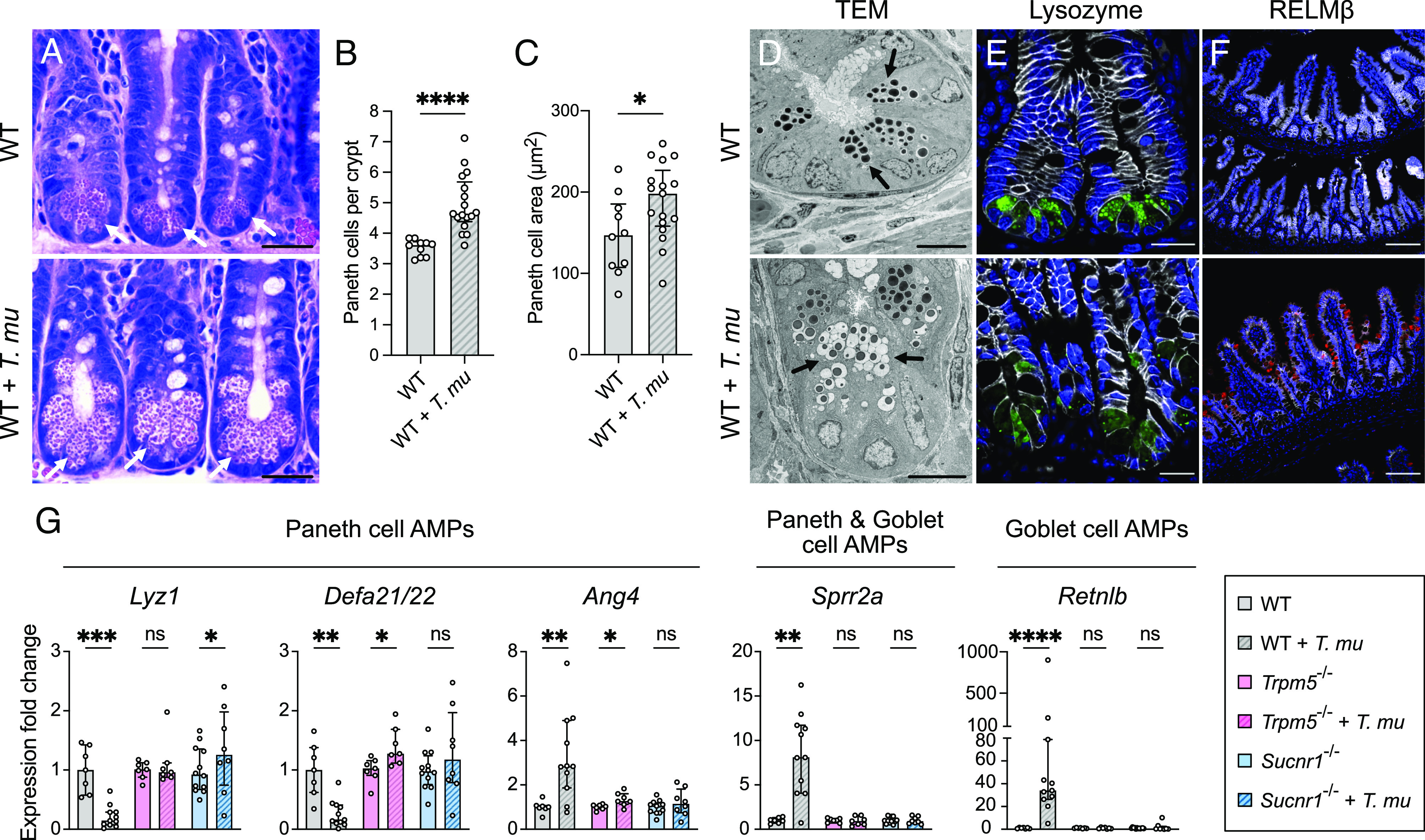
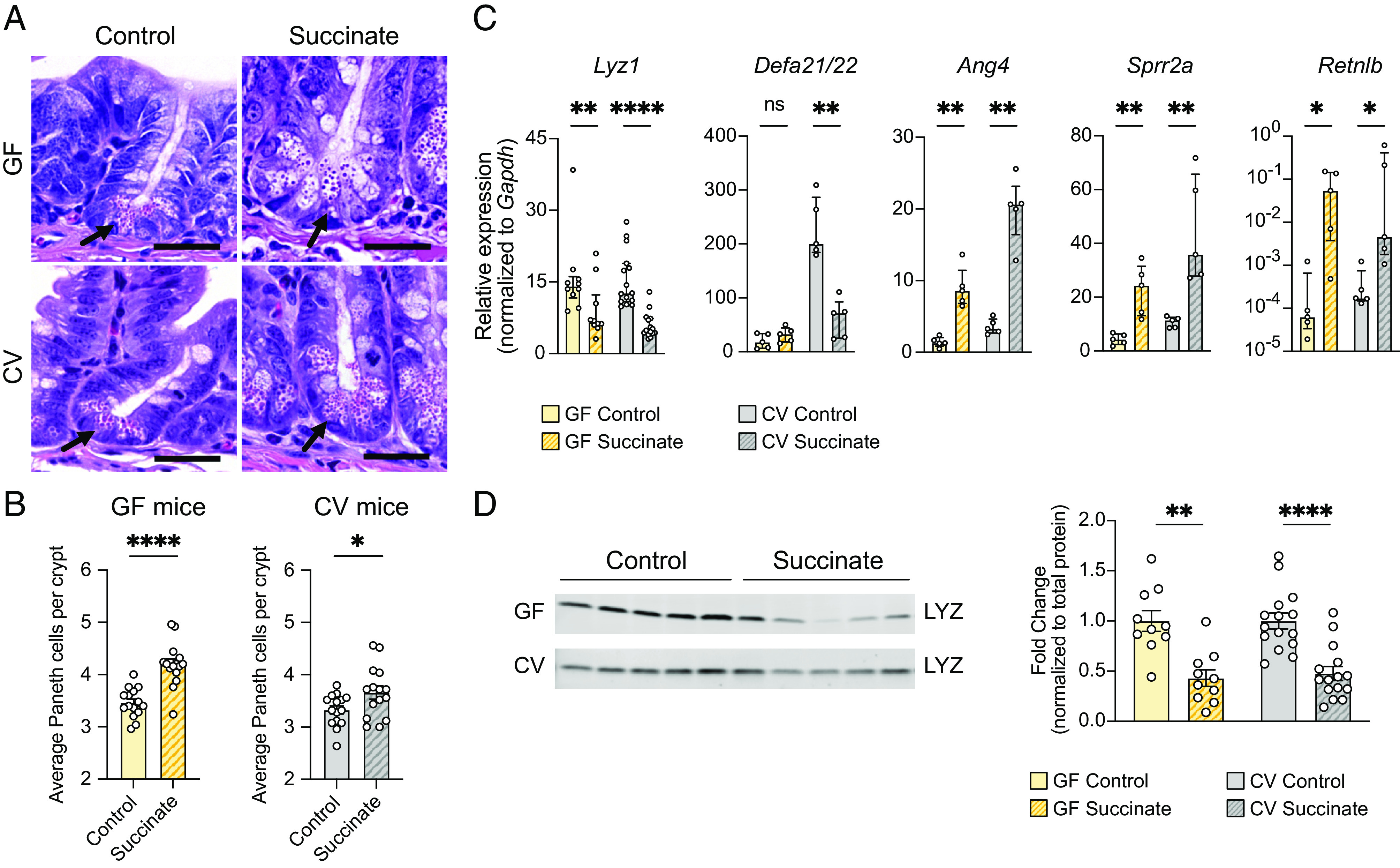
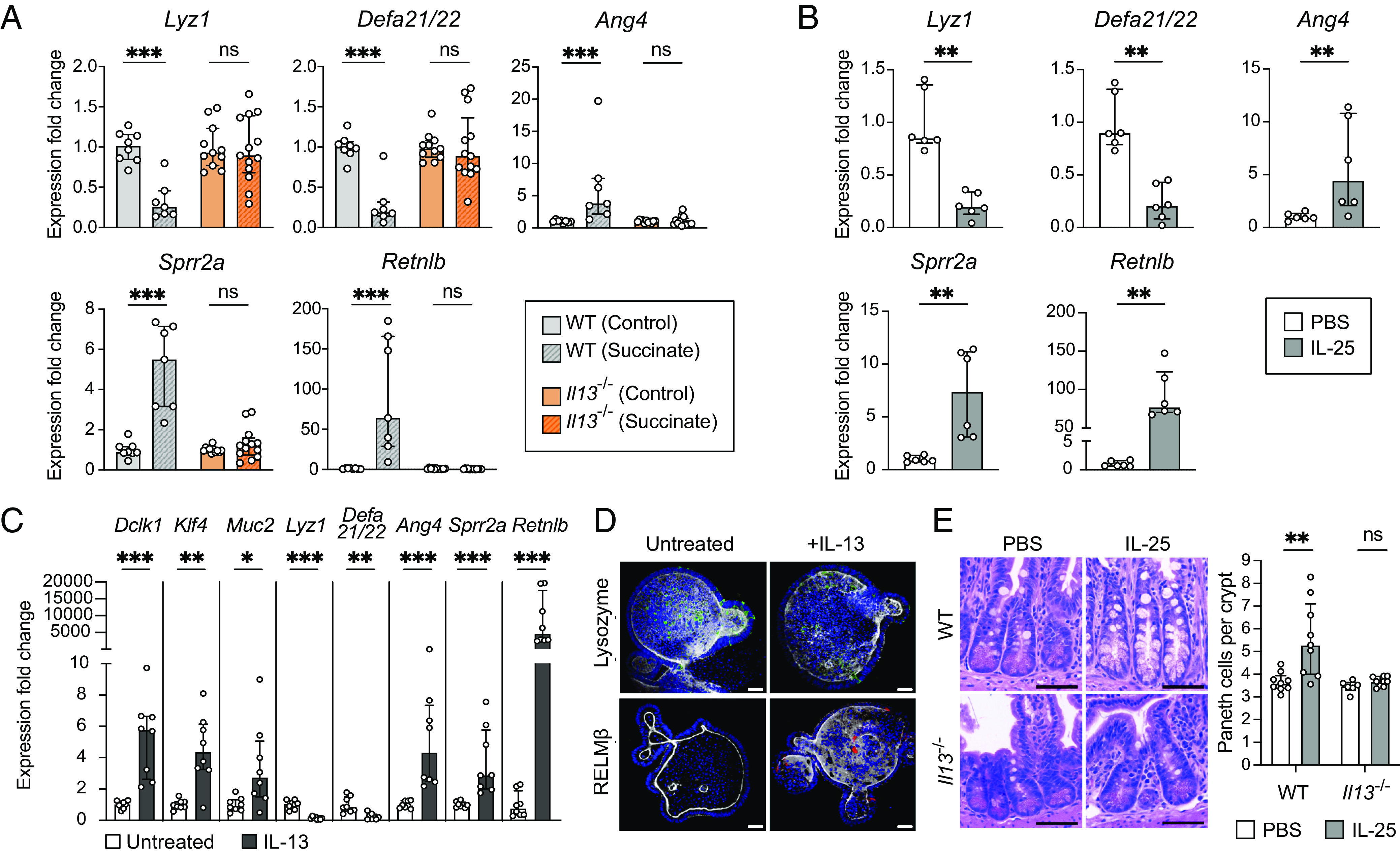
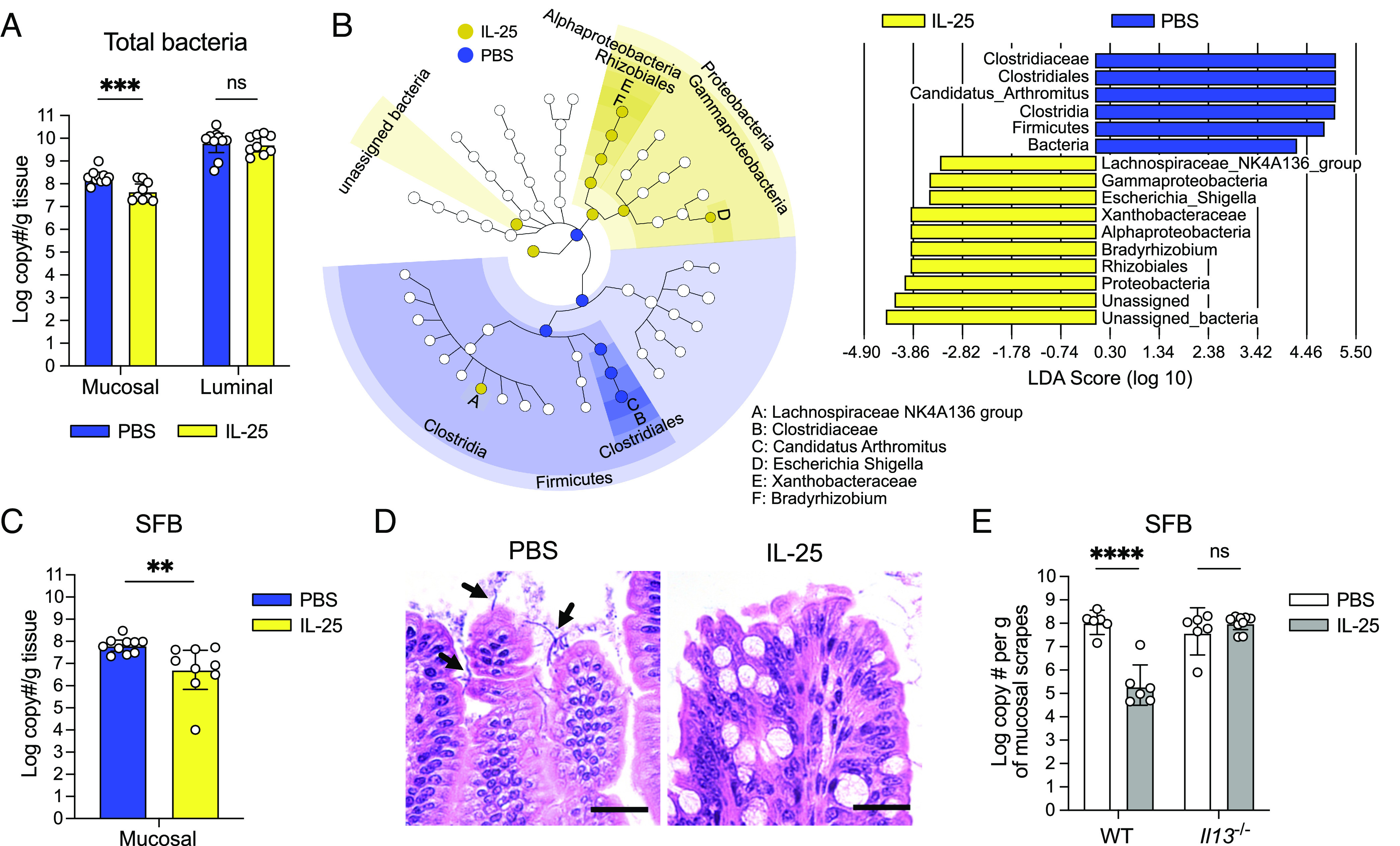
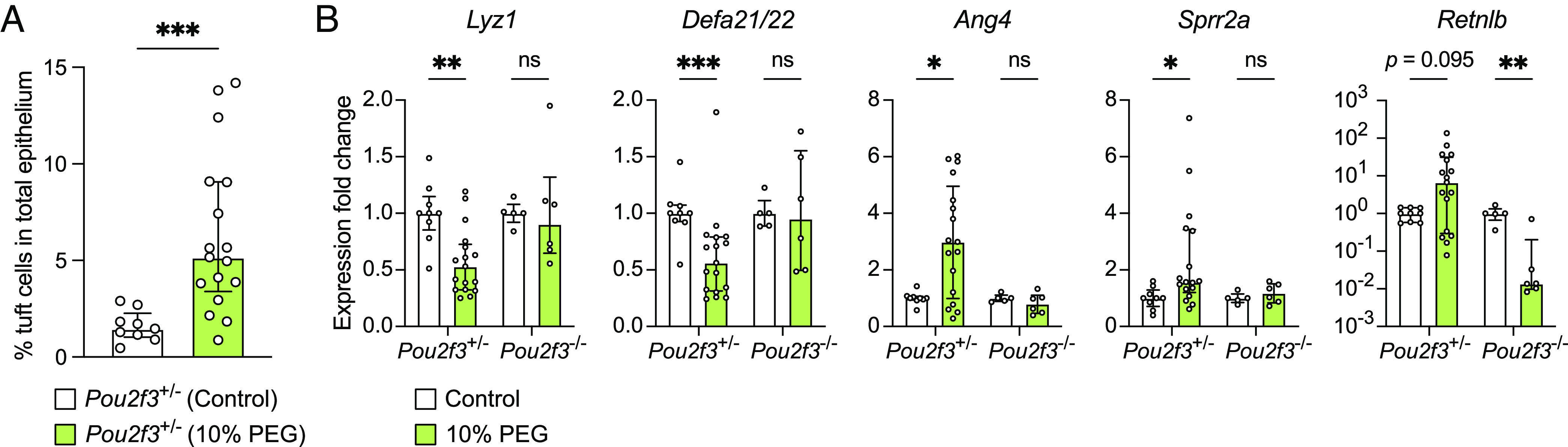
Succinate produced by the commensal protist Tritrichomonas musculis (T. mu) stimulates chemosensory tuft cells, resulting in intestinal type 2 immunity. Tuft cells express the succinate receptor SUCNR1, yet this receptor does not mediate antihelminth immunity nor alter protist colonization. Here, we report that microbial-derived succinate increases Paneth cell numbers and profoundly alters the antimicrobial peptide (AMP) landscape in the small intestine. Succinate was sufficient to drive this epithelial remodeling, but not in mice lacking tuft cell chemosensory components required to detect this metabolite. Tuft cells respond to succinate by stimulating type 2 immunity, leading to interleukin-13-mediated epithelial and AMP expression changes. Moreover, type 2 immunity decreases the total number of mucosa-associated bacteria and alters the small intestinal microbiota composition. Finally, tuft cells can detect short-term bacterial dysbiosis that leads to a spike in luminal succinate levels and modulate AMP production in response. These findings demonstrate that a single metabolite produced by commensals can markedly shift the intestinal AMP profile and suggest that tuft cells utilize SUCNR1 and succinate sensing to modulate bacterial homeostasis.
Keywords: Paneth cell; antimicrobial peptides; succinate; tuft cell; type 2 immunity.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
