

Acute downregulation of emerin alters actomyosin cytoskeleton connectivity and function
- PMID: 37254483
- PMCID: PMC10541481
- DOI: 10.1016/j.bpj.2023.05.027
Acute downregulation of emerin alters actomyosin cytoskeleton connectivity and function
Abstract
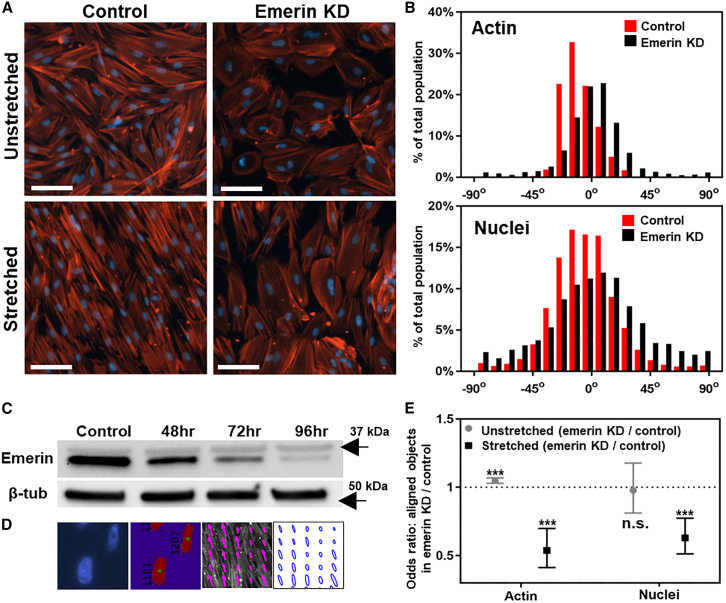
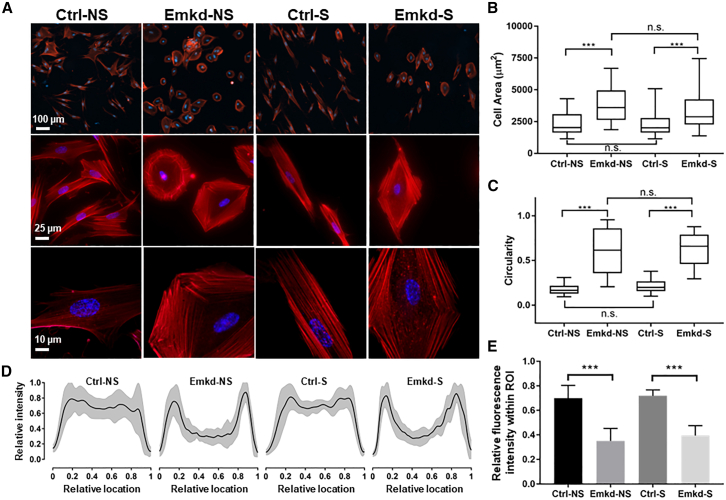
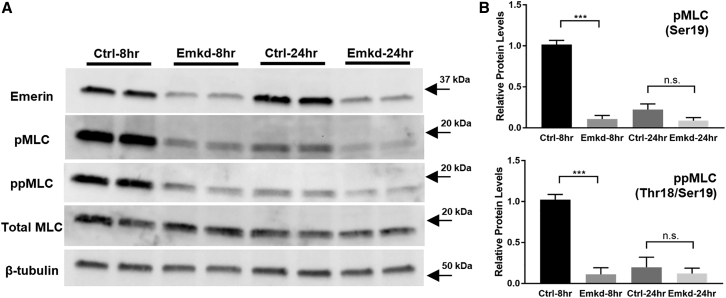
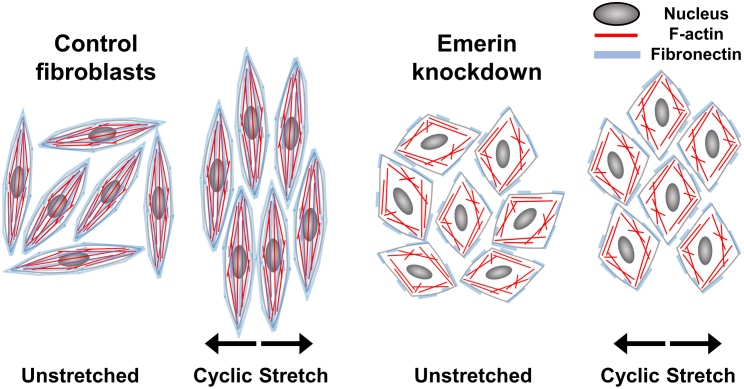
Fetal lung fibroblasts contribute dynamic infrastructure for the developing lung. These cells undergo dynamic mechanical transitions, including cyclic stretch and spreading, which are integral to lung growth in utero. We investigated the role of the nuclear envelope protein emerin in cellular responses to these dynamic mechanical transitions. In contrast to control cells, which briskly realigned their nuclei, actin cytoskeleton, and extracellular matrices in response to cyclic stretch, fibroblasts that were acutely downregulated for emerin showed incomplete reorientation of both nuclei and actin cytoskeleton. Emerin-downregulated fibroblasts were also aberrantly circular in contrast to the spindle-shaped controls and exhibited an altered pattern of filamentous actin organization that was disconnected from the nucleus. Emerin knockdown was also associated with reduced myosin light chain phosphorylation during cell spreading. Interestingly, emerin-downregulated fibroblasts also demonstrated reduced fibronectin fibrillogenesis and production. These findings indicate that nuclear-cytoskeletal coupling serves a role in the dynamic regulation of cytoskeletal structure and function and may also impact the transmission of traction force to the extracellular matrix microenvironment.
Copyright © 2023 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures


References
-
- De Vries J.I., Visser G.H., Prechtl H.F. The emergence of fetal behaviour. II. Quantitative aspects. Early Hum. Dev. 1985;12:99–120. - PubMed
-
- Natale R., Nasello-Paterson C., Connors G. Patterns of fetal breathing activity in the human fetus at 24 to 28 weeks of gestation. Am. J. Obstet. Gynecol. 1988;158:317–321. - PubMed
-
- Patrick J., Campbell K., et al. Richardson B. Patterns of human fetal breathing during the last 10 weeks of pregnancy. Obstet. Gynecol. 1980;56:24–30. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
