

PSMC3 proteasome subunit variants are associated with neurodevelopmental delay and type I interferon production
- PMID: 37256937
- PMCID: PMC10506367
- DOI: 10.1126/scitranslmed.abo3189
PSMC3 proteasome subunit variants are associated with neurodevelopmental delay and type I interferon production
Abstract
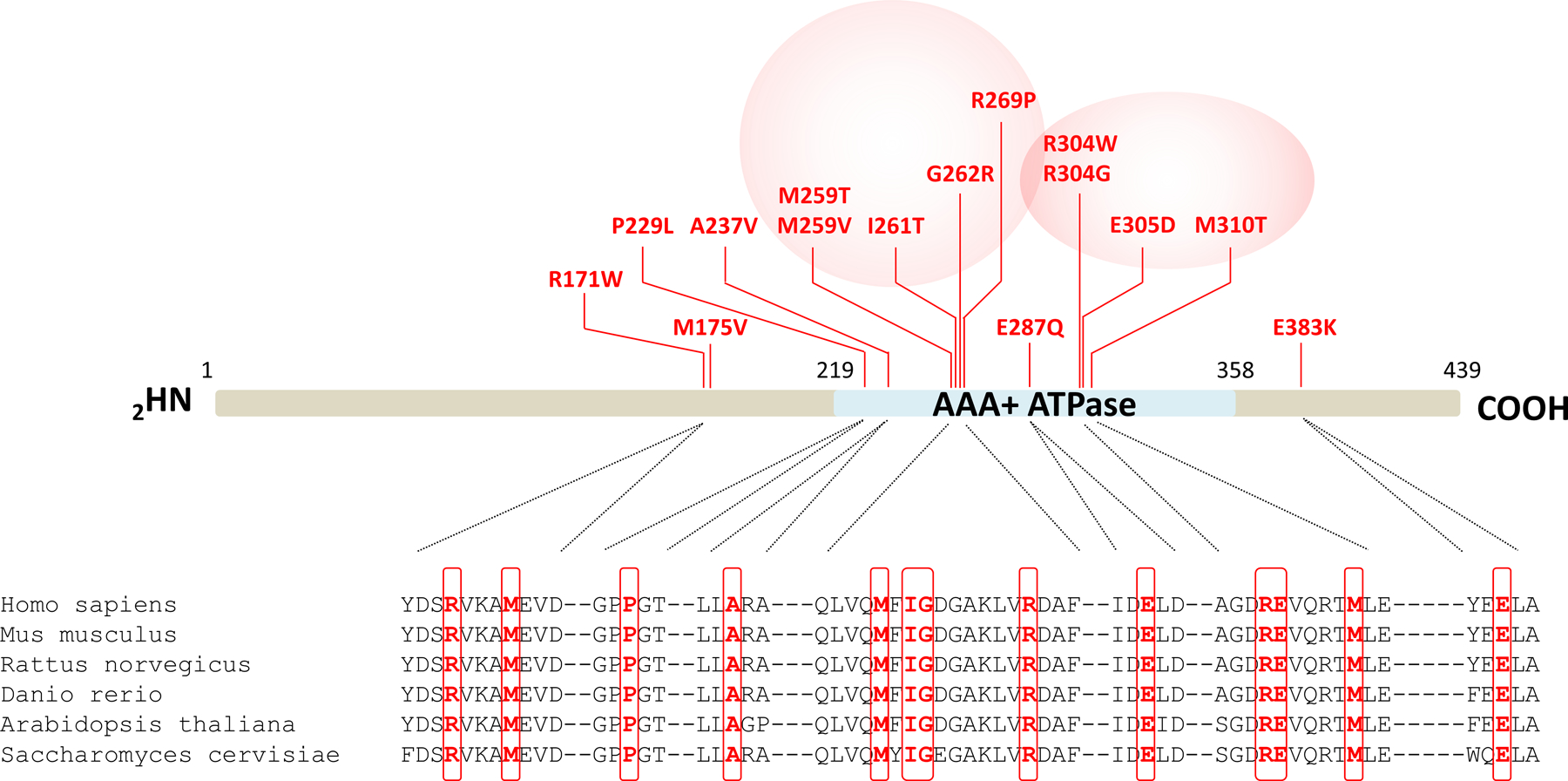
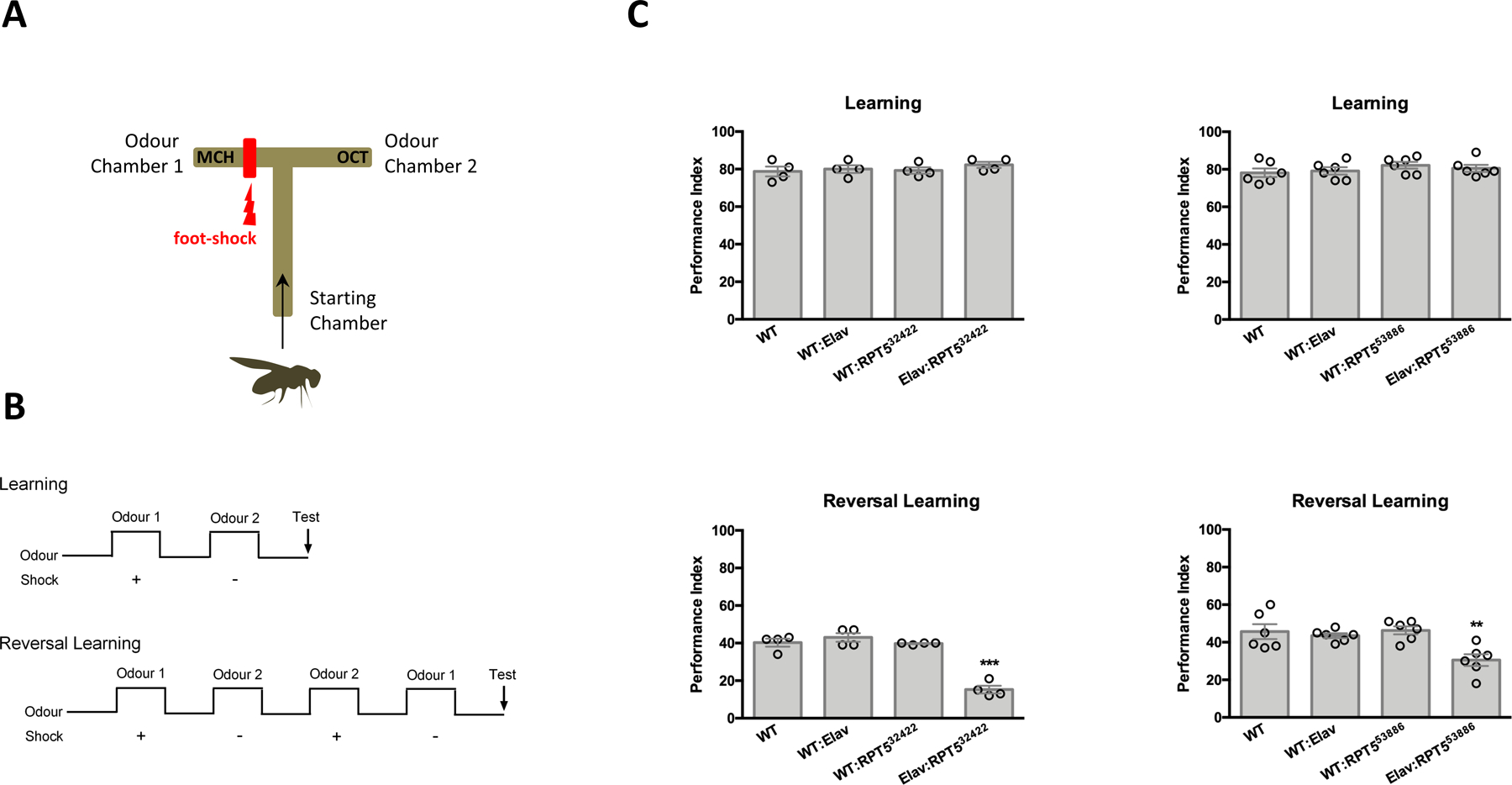
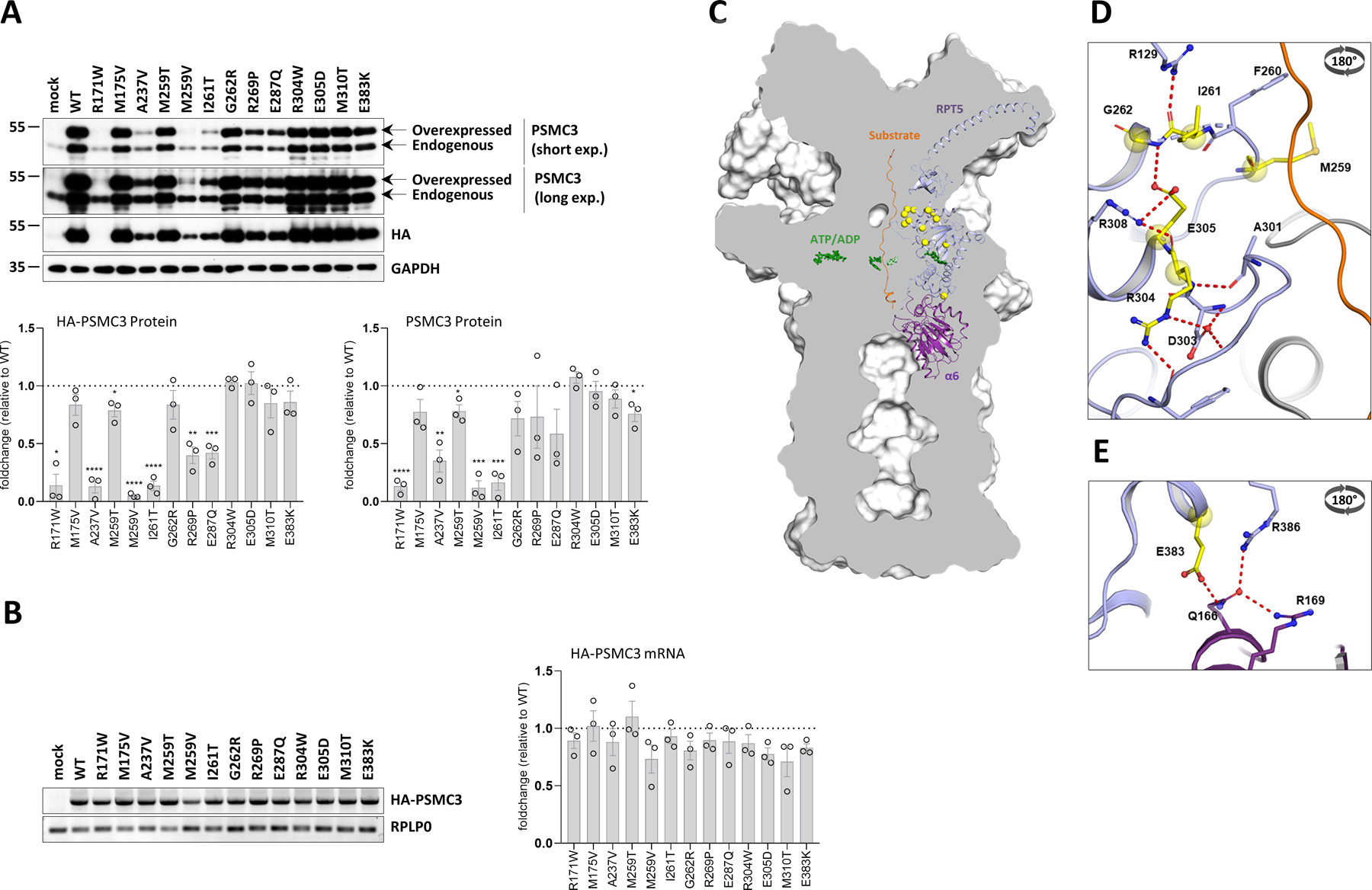
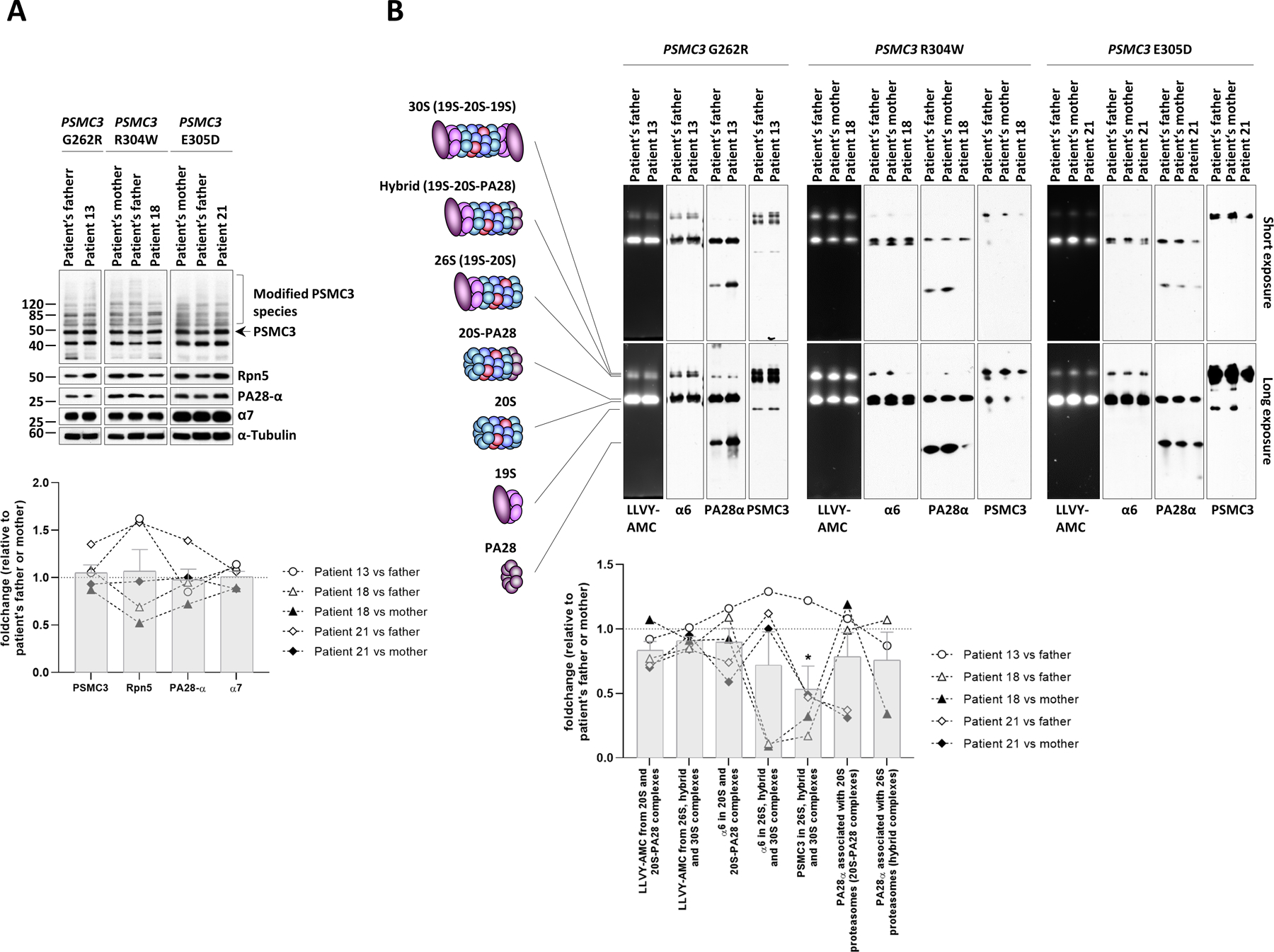
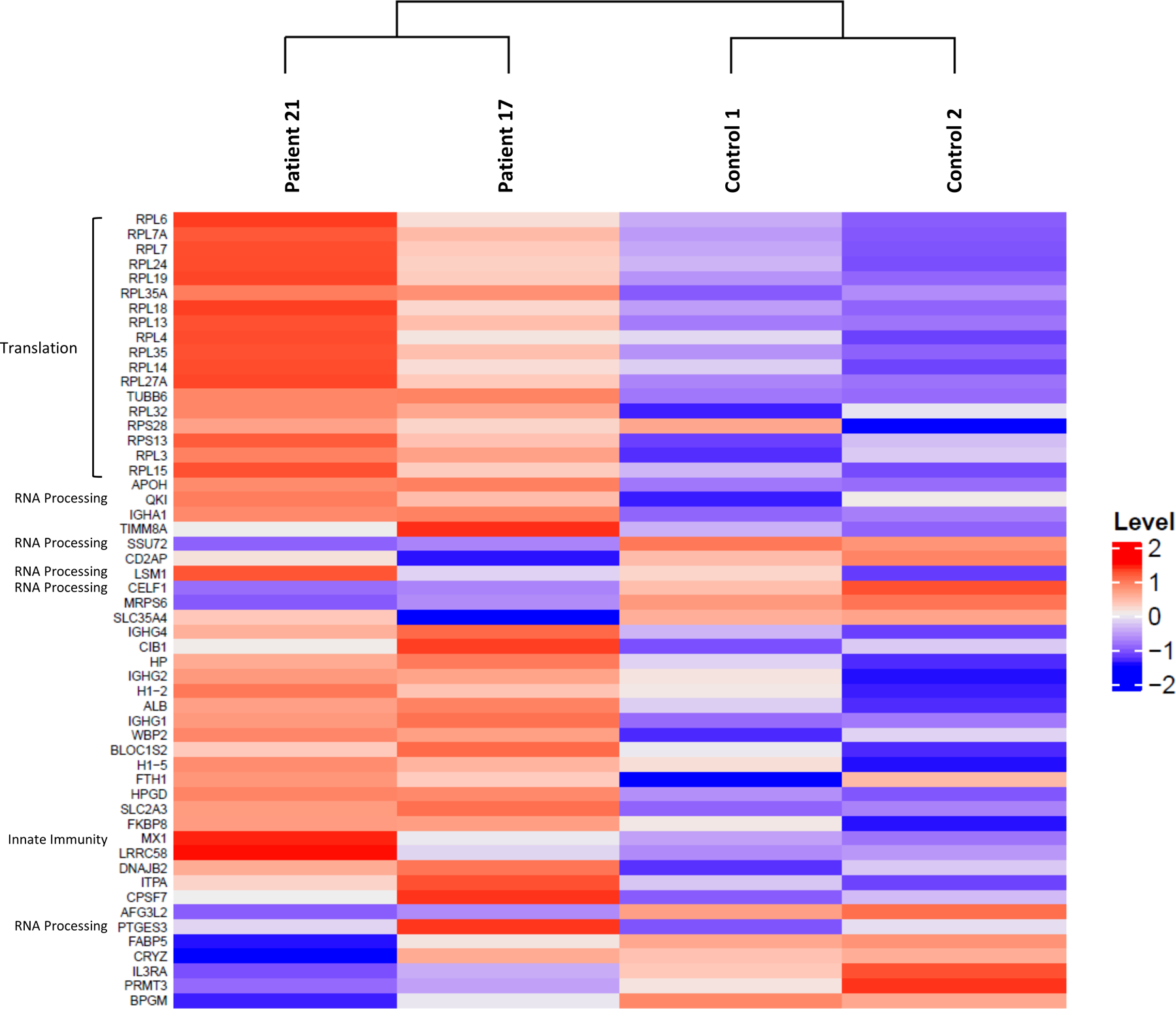
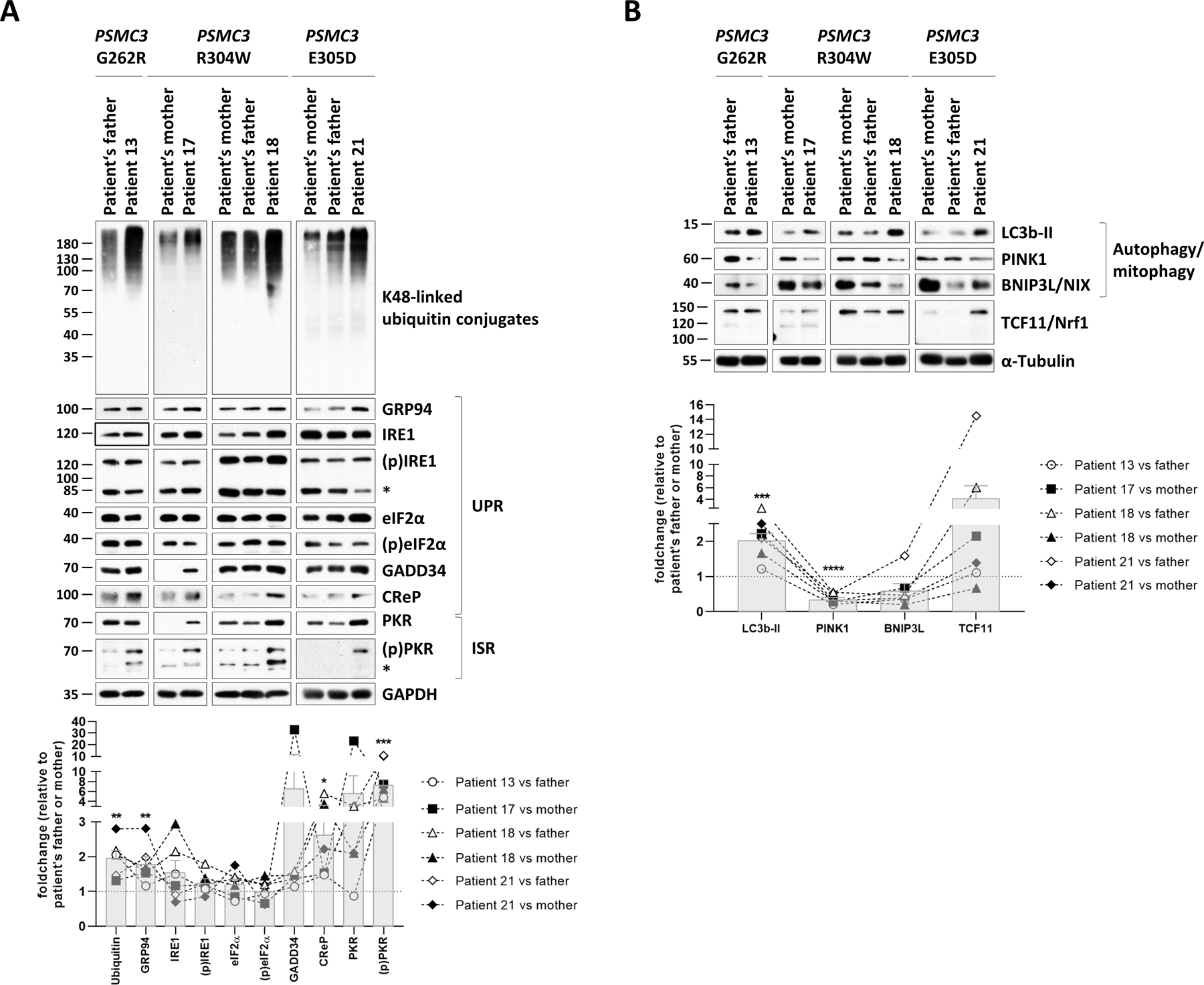
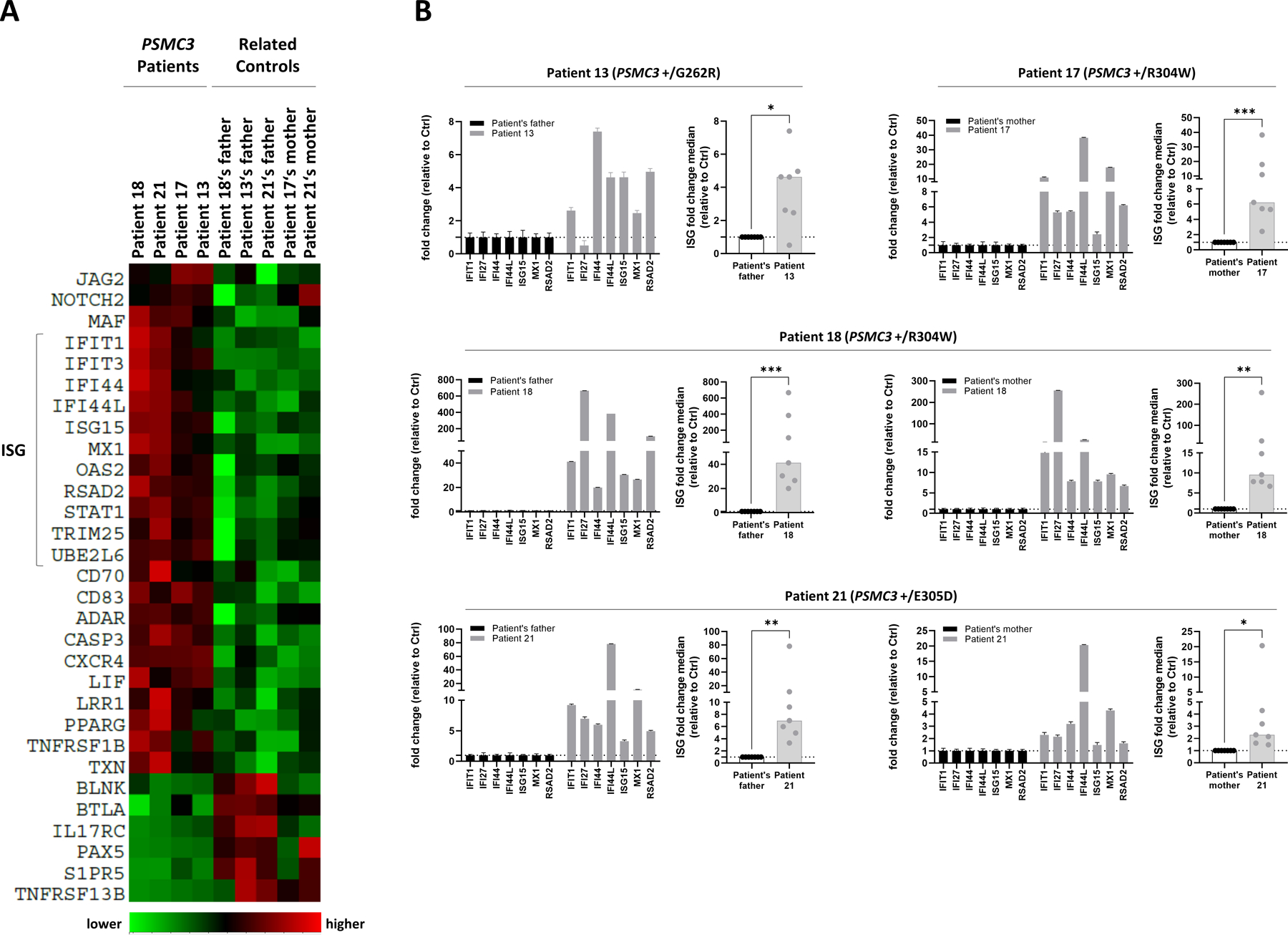
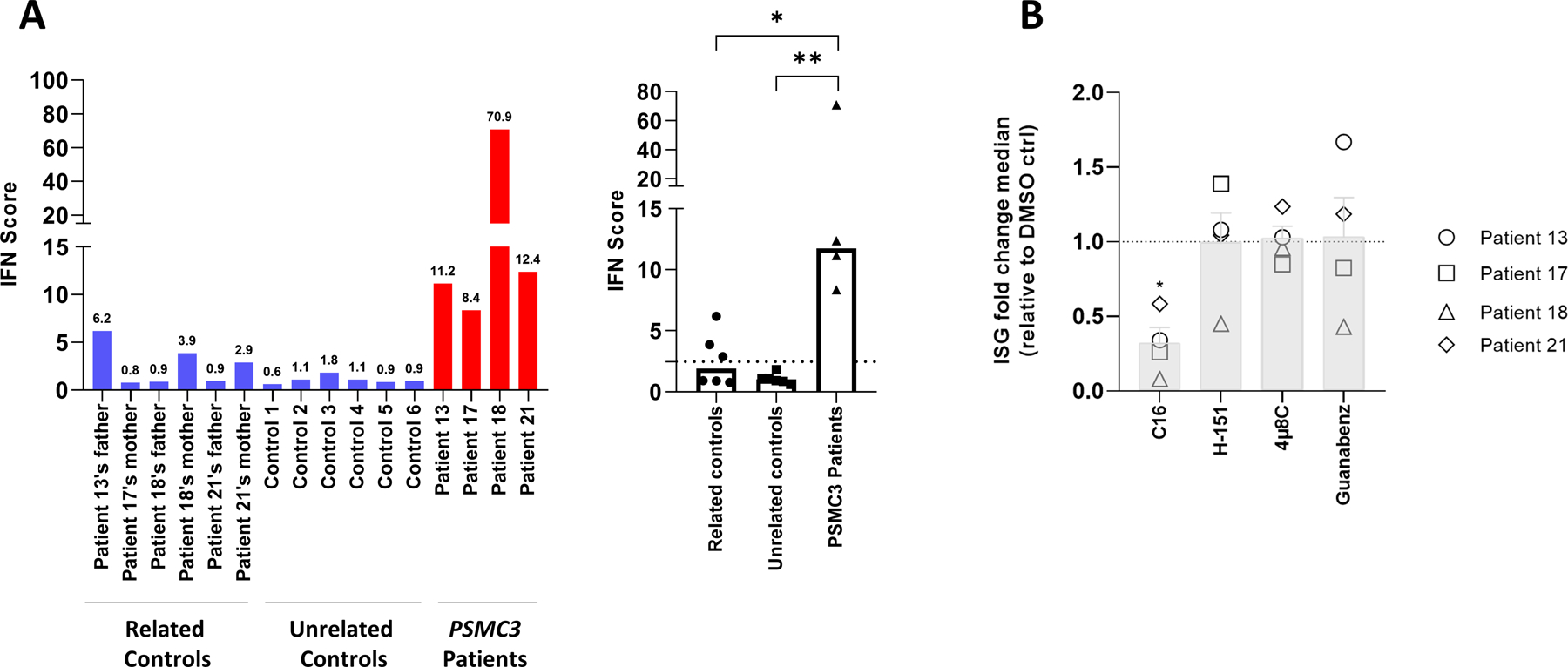
A critical step in preserving protein homeostasis is the recognition, binding, unfolding, and translocation of protein substrates by six AAA-ATPase proteasome subunits (ATPase-associated with various cellular activities) termed PSMC1-6, which are required for degradation of proteins by 26S proteasomes. Here, we identified 15 de novo missense variants in the PSMC3 gene encoding the AAA-ATPase proteasome subunit PSMC3/Rpt5 in 23 unrelated heterozygous patients with an autosomal dominant form of neurodevelopmental delay and intellectual disability. Expression of PSMC3 variants in mouse neuronal cultures led to altered dendrite development, and deletion of the PSMC3 fly ortholog Rpt5 impaired reversal learning capabilities in fruit flies. Structural modeling as well as proteomic and transcriptomic analyses of T cells derived from patients with PSMC3 variants implicated the PSMC3 variants in proteasome dysfunction through disruption of substrate translocation, induction of proteotoxic stress, and alterations in proteins controlling developmental and innate immune programs. The proteostatic perturbations in T cells from patients with PSMC3 variants correlated with a dysregulation in type I interferon (IFN) signaling in these T cells, which could be blocked by inhibition of the intracellular stress sensor protein kinase R (PKR). These results suggest that proteotoxic stress activated PKR in patient-derived T cells, resulting in a type I IFN response. The potential relationship among proteosome dysfunction, type I IFN production, and neurodevelopment suggests new directions in our understanding of pathogenesis in some neurodevelopmental disorders.
Conflict of interest statement
Figures
References
-
- Tanaka K, Mizushima T, Saeki Y. The proteasome: molecular machinery and pathophysiological roles. Biol Chem 393, 217–34 (2012). - PubMed
-
- Goloubinoff P. Mechanisms of protein homeostasis in health, aging and disease. Swiss Med Wkly 146, w14306 (2016). - PubMed
-
- Seifert U, Bialy LP, Ebstein F, Bech-Otschir D, Voigt A, Schröter F, Prozorovski T, Lange N, Steffen J, Rieger M, Kuckelkorn U, Aktas O, Kloetzel P-M, Krüger E. Immunoproteasomes preserve protein homeostasis upon interferon-induced oxidative stress. Cell 142, 613–24 (2010). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
