

Germ cell-specific eIF4E1b regulates maternal mRNA translation to ensure zygotic genome activation
- PMID: 37257918
- PMCID: PMC10270193
- DOI: 10.1101/gad.350400.123
Germ cell-specific eIF4E1b regulates maternal mRNA translation to ensure zygotic genome activation
Abstract
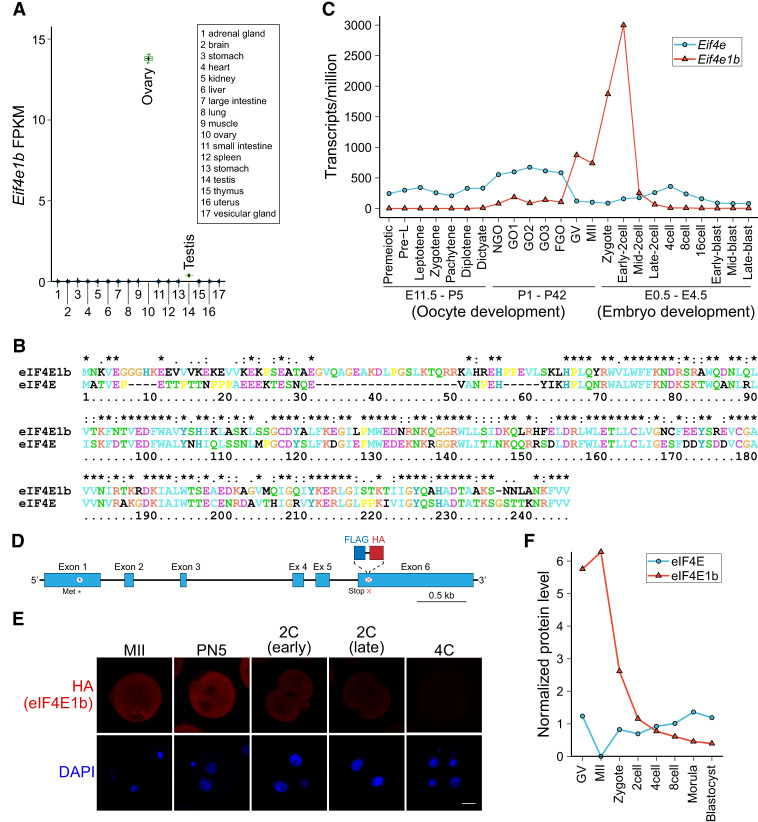
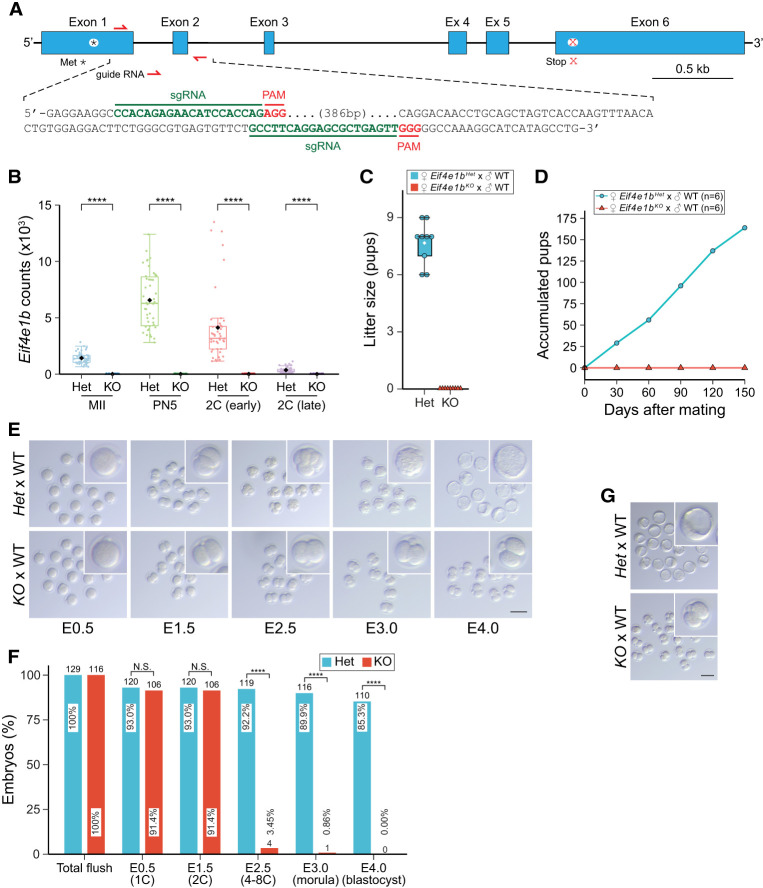
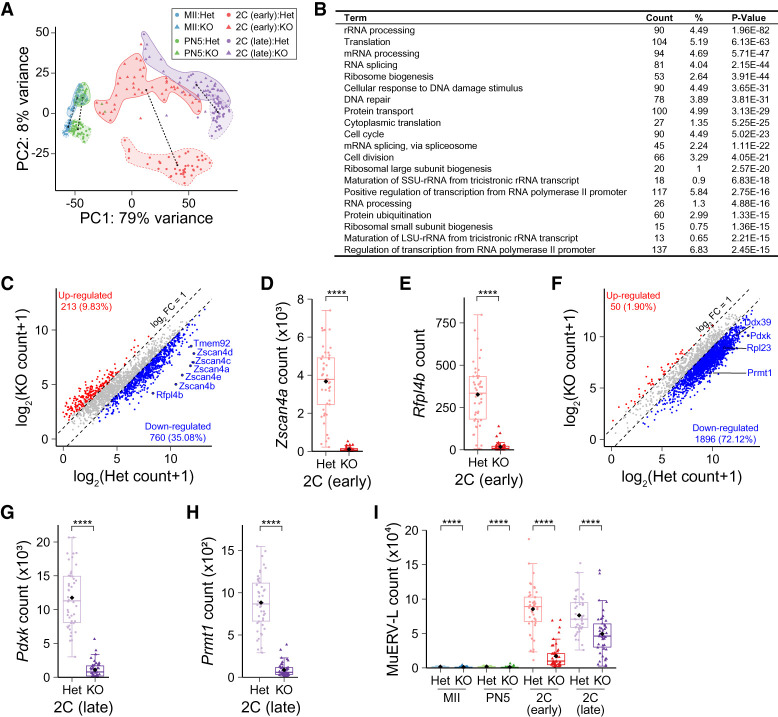
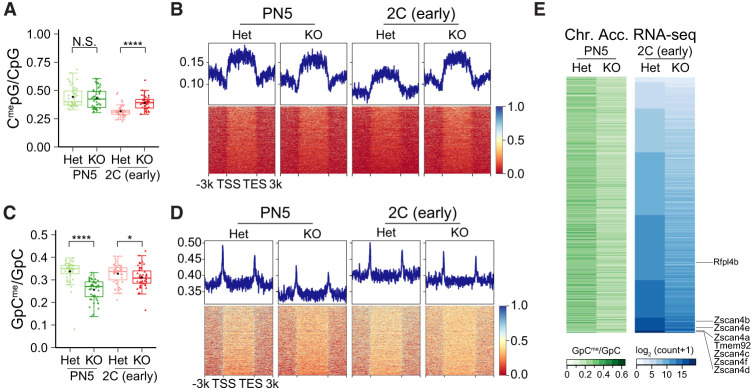
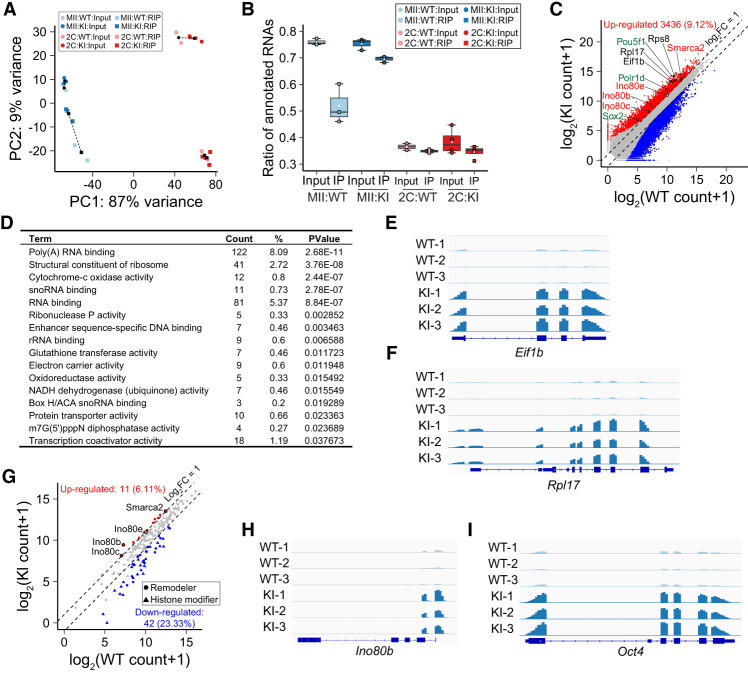
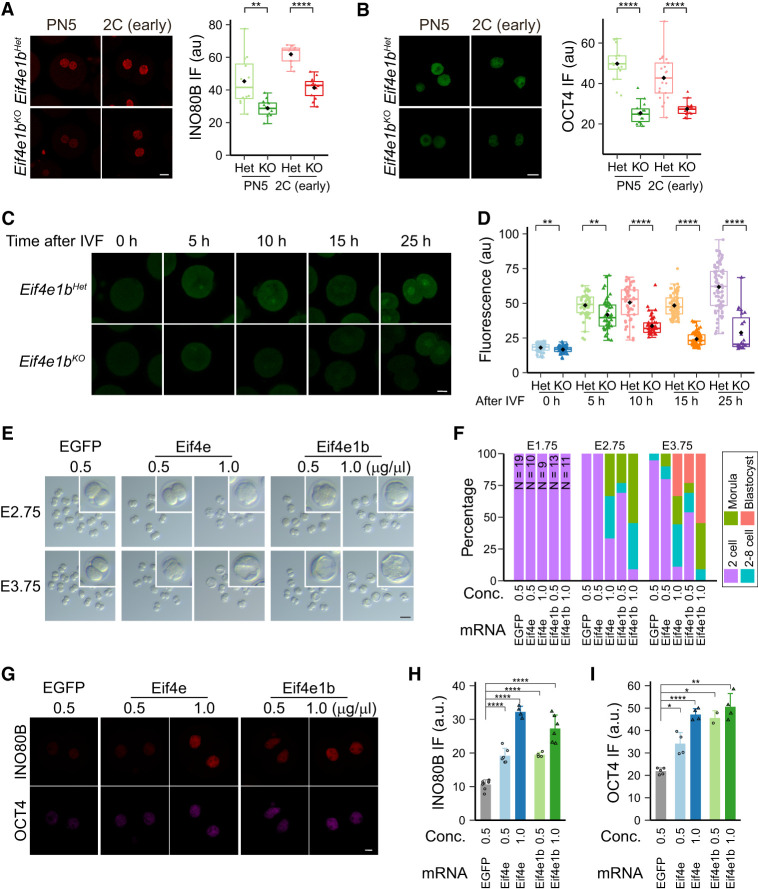
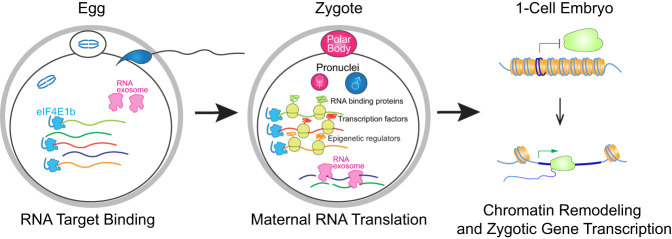
Translation of maternal mRNAs is detected before transcription of zygotic genes and is essential for mammalian embryo development. How certain maternal mRNAs are selected for translation instead of degradation and how this burst of translation affects zygotic genome activation remain unknown. Using gene-edited mice, we document that the oocyte-specific eukaryotic translation initiation factor 4E family member 1b (eIF4E1b) is the regulator of maternal mRNA expression that ensures subsequent reprogramming of the zygotic genome. In oocytes, eIF4E1b binds to transcripts encoding translation machinery proteins, chromatin remodelers, and reprogramming factors to promote their translation in zygotes and protect them from degradation. The protein products are thought to establish an open chromatin landscape in one-cell zygotes to enable transcription of genes required for cleavage stage development. Our results define a program for rapid resetting of the zygotic epigenome that is regulated by maternal mRNA expression and provide new insights into the mammalian maternal-to-zygotic transition.
Keywords: Eif4e1b; maternal RNA translation; mouse embryo; zygotic genome activation.
Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Alda-Catalinas C, Bredikhin D, Hernando-Herraez I, Santos F, Kubinyecz O, Eckersley-Maslin MA, Stegle O, Reik W. 2020. A single-cell transcriptomics CRISPR-activation screen identifies epigenetic regulators of the zygotic genome activation program. Cell Syst 11: 25–41.e9. 10.1016/j.cels.2020.06.004 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases