

Interleukin 1β triggers synaptic and memory deficits in Herpes simplex virus type-1-infected mice by downregulating the expression of synaptic plasticity-related genes via the epigenetic MeCP2/HDAC4 complex
- PMID: 37261502
- PMCID: PMC10234878
- DOI: 10.1007/s00018-023-04817-5
Interleukin 1β triggers synaptic and memory deficits in Herpes simplex virus type-1-infected mice by downregulating the expression of synaptic plasticity-related genes via the epigenetic MeCP2/HDAC4 complex
Abstract
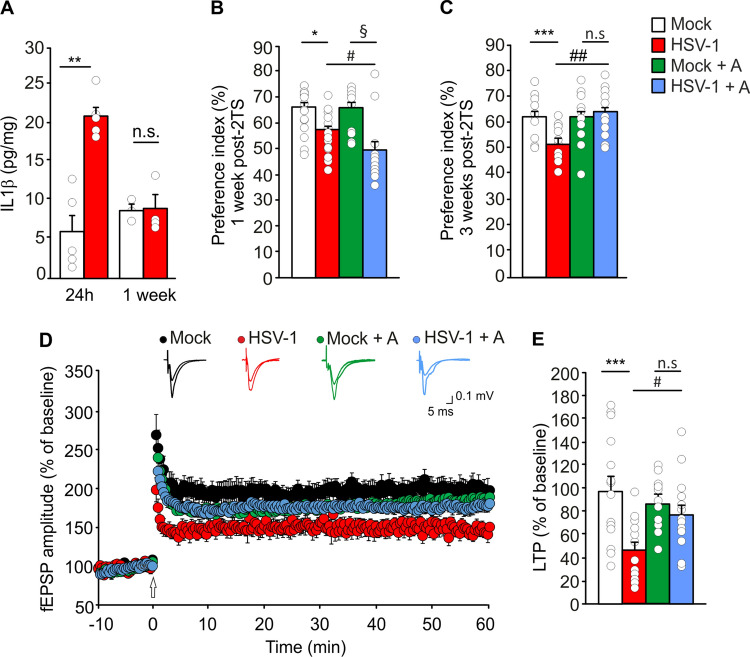
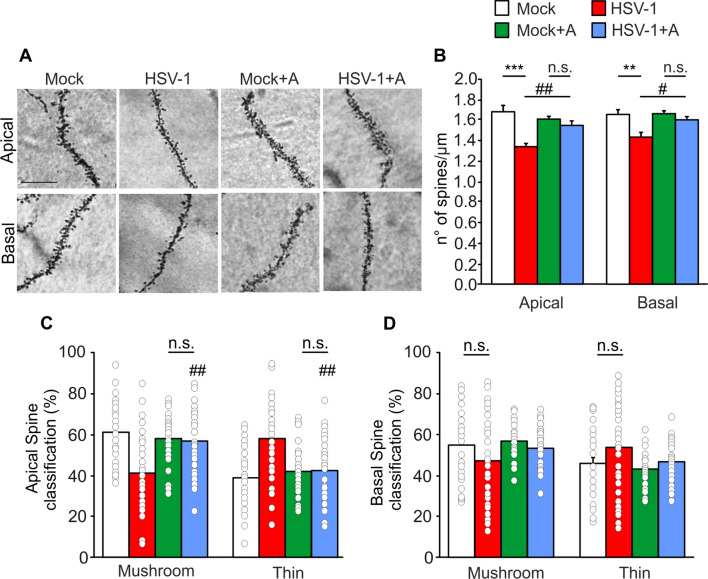
Extensive research provides evidence that neuroinflammation underlies numerous brain disorders. However, the molecular mechanisms by which inflammatory mediators determine synaptic and cognitive dysfunction occurring in neurodegenerative diseases (e.g., Alzheimer's disease) are far from being fully understood. Here we investigated the role of interleukin 1β (IL-1β), and the molecular cascade downstream the activation of its receptor, to the synaptic dysfunction occurring in the mouse model of multiple Herpes simplex virus type-1 (HSV-1) reactivations within the brain. These mice are characterized by neuroinflammation and memory deficits associated with a progressive accumulation of neurodegenerative hallmarks (e.g., amyloid-β protein and tau hyperphosphorylation). Here we show that mice undergone two HSV-1 reactivations in the brain exhibited increased levels of IL-1β along with significant alterations of: (1) cognitive performances; (2) hippocampal long-term potentiation; (3) expression synaptic-related genes and pre- and post-synaptic proteins; (4) dendritic spine density and morphology. These effects correlated with activation of the epigenetic repressor MeCP2 that, in association with HDAC4, affected the expression of synaptic plasticity-related genes. Specifically, in response to HSV-1 infection, HDAC4 accumulated in the nucleus and promoted MeCP2 SUMOylation that is a post-translational modification critically affecting the repressive activity of MeCP2. The blockade of IL-1 receptors by the specific antagonist Anakinra prevented the MeCP2 increase and the consequent downregulation of gene expression along with rescuing structural and functional indices of neurodegeneration. Collectively, our findings provide novel mechanistic evidence on the role played by HSV-1-activated IL-1β signaling pathways in synaptic deficits leading to cognitive impairment.
Keywords: Anakinra; Memory; Neurodegeneration; Neuroinflammation; SUMOylation; Synaptic function.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing financial interests.
Figures



Similar articles
-
Recurrent herpes simplex virus-1 infection induces hallmarks of neurodegeneration and cognitive deficits in mice.PLoS Pathog. 2019 Mar 14;15(3):e1007617. doi: 10.1371/journal.ppat.1007617. eCollection 2019 Mar. PLoS Pathog. 2019. PMID: 30870531 Free PMC article.
-
MeCP2 prevents age-associated cognitive decline via restoring synaptic plasticity in a senescence-accelerated mouse model.Aging Cell. 2021 Sep;20(9):e13451. doi: 10.1111/acel.13451. Epub 2021 Aug 7. Aging Cell. 2021. PMID: 34363729 Free PMC article.
-
Targeting SIK3 to modulate hippocampal synaptic plasticity and cognitive function by regulating the transcription of HDAC4 in a mouse model of Alzheimer's disease.Neuropsychopharmacology. 2024 May;49(6):942-952. doi: 10.1038/s41386-023-01775-1. Epub 2023 Dec 6. Neuropsychopharmacology. 2024. PMID: 38057370 Free PMC article.
-
Synaptic Plasticity, Dementia and Alzheimer Disease.CNS Neurol Disord Drug Targets. 2017;16(3):220-233. doi: 10.2174/1871527316666170113120853. CNS Neurol Disord Drug Targets. 2017. PMID: 28088900 Review.
-
Epigenetic Mechanisms in Memory and Cognitive Decline Associated with Aging and Alzheimer's Disease.Int J Mol Sci. 2021 Nov 13;22(22):12280. doi: 10.3390/ijms222212280. Int J Mol Sci. 2021. PMID: 34830163 Free PMC article. Review.
Cited by
-
Estrogen-immuno-neuromodulation disorders in menopausal depression.J Neuroinflammation. 2024 Jun 19;21(1):159. doi: 10.1186/s12974-024-03152-1. J Neuroinflammation. 2024. PMID: 38898454 Free PMC article. Review.
-
Regulation of inflammatory cytokines and activation of PI3K/Akt pathway by Yiqi Jiedu Formula in recurrent Herpes Simplex Keratitis: Experimental and network pharmacology evidence.Virus Res. 2025 May;355:199561. doi: 10.1016/j.virusres.2025.199561. Epub 2025 Mar 20. Virus Res. 2025. PMID: 40120648 Free PMC article.
-
A novel approach to metabolic profiling in case models of MECP2-related disorders.Metab Brain Dis. 2025 Feb 13;40(2):124. doi: 10.1007/s11011-025-01546-5. Metab Brain Dis. 2025. PMID: 39945871 Free PMC article.
-
Unveiling the Involvement of Herpes Simplex Virus-1 in Alzheimer's Disease: Possible Mechanisms and Therapeutic Implications.Mol Neurobiol. 2025 May;62(5):5850-5874. doi: 10.1007/s12035-024-04535-4. Epub 2024 Dec 9. Mol Neurobiol. 2025. PMID: 39648189 Review.
-
Dysmaturation of sleep state and electroencephalographic activity after hypoxia-ischaemia in preterm fetal sheep.J Cereb Blood Flow Metab. 2024 Aug;44(8):1376-1392. doi: 10.1177/0271678X241236014. Epub 2024 Feb 28. J Cereb Blood Flow Metab. 2024. PMID: 38415649 Free PMC article.
References
-
- Cacabelos R, Alvarez XA, Fernández-Novoa L, et al. Brain interleukin-1 beta in Alzheimer’s disease and vascular dementia. Methods Find Exp Clin Pharmacol. 1994;16:141–151. - PubMed
MeSH terms
Substances
Grants and funding
- 2017A9MK4R_004/Ministero dell'Istruzione, dell'Università e della Ricerca
- FOE2020:project title "Nuovi biomarker diagnostici e terapeutici delle malattie neurodegenerative"/Ministero dell'Istruzione, dell'Università e della Ricerca
- 20179JHAMZ_006/Ministero dell'Istruzione, dell'Università e della Ricerca
- Ricerca Corrente 2023 - Fondazione Policlinico Universitario A. Gemelli IRCCS/Ministero della Salute
- D1 intramural funds/Università Cattolica del Sacro Cuore
LinkOut - more resources
Full Text Sources
Medical
