

Liver and muscle circadian clocks cooperate to support glucose tolerance in mice
- PMID: 37267101
- PMCID: PMC10592114
- DOI: 10.1016/j.celrep.2023.112588
Liver and muscle circadian clocks cooperate to support glucose tolerance in mice
Abstract
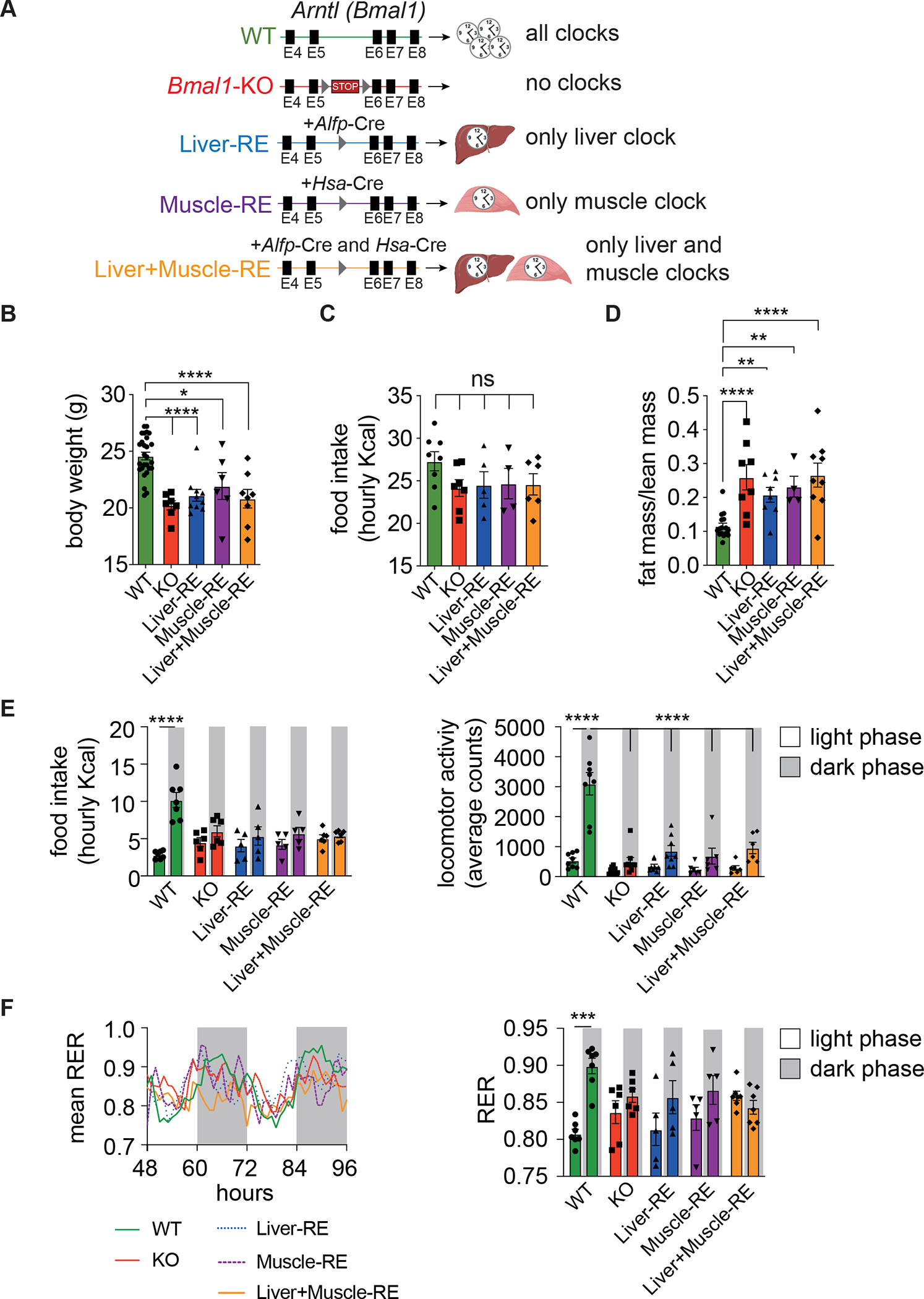
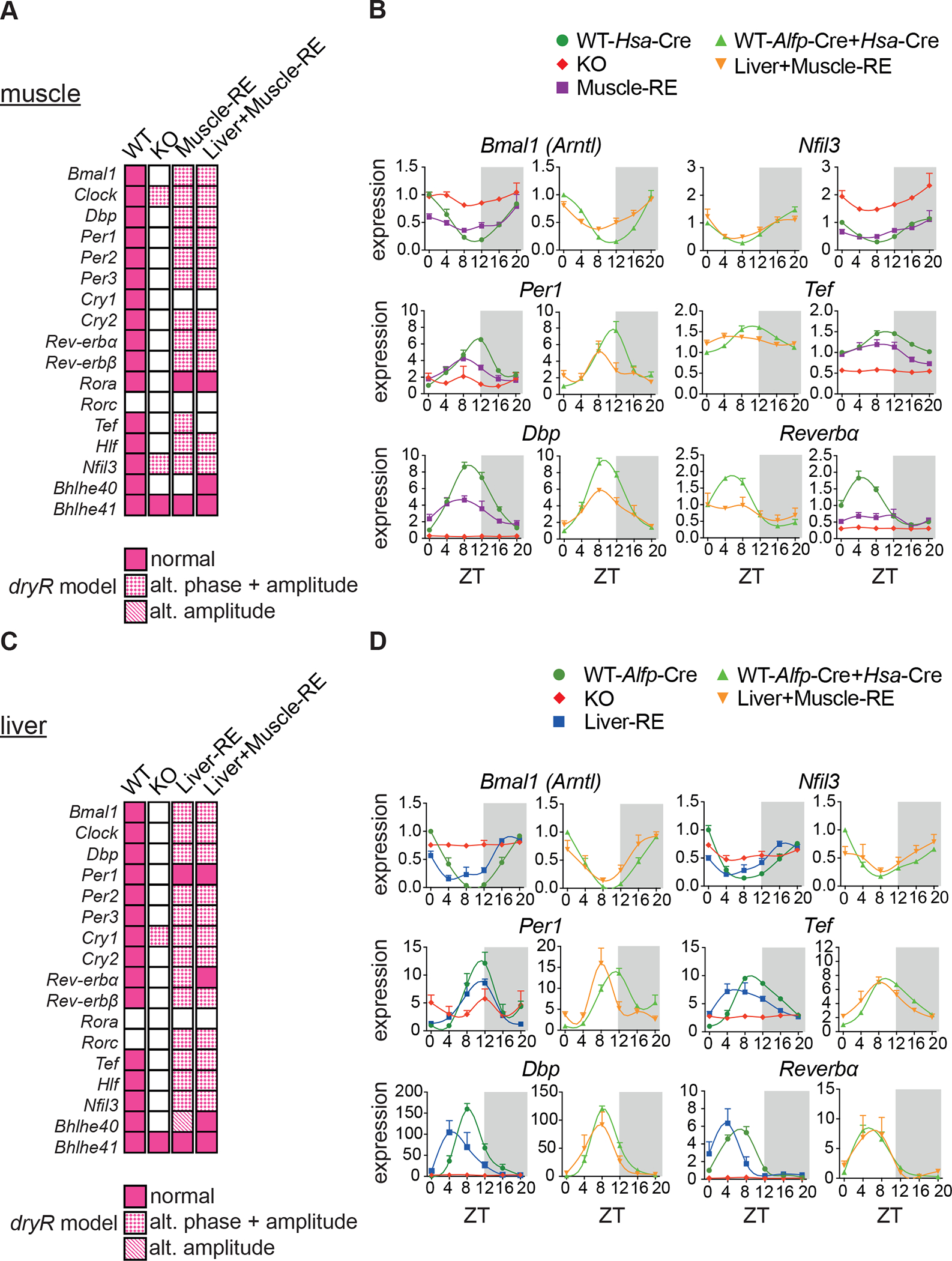
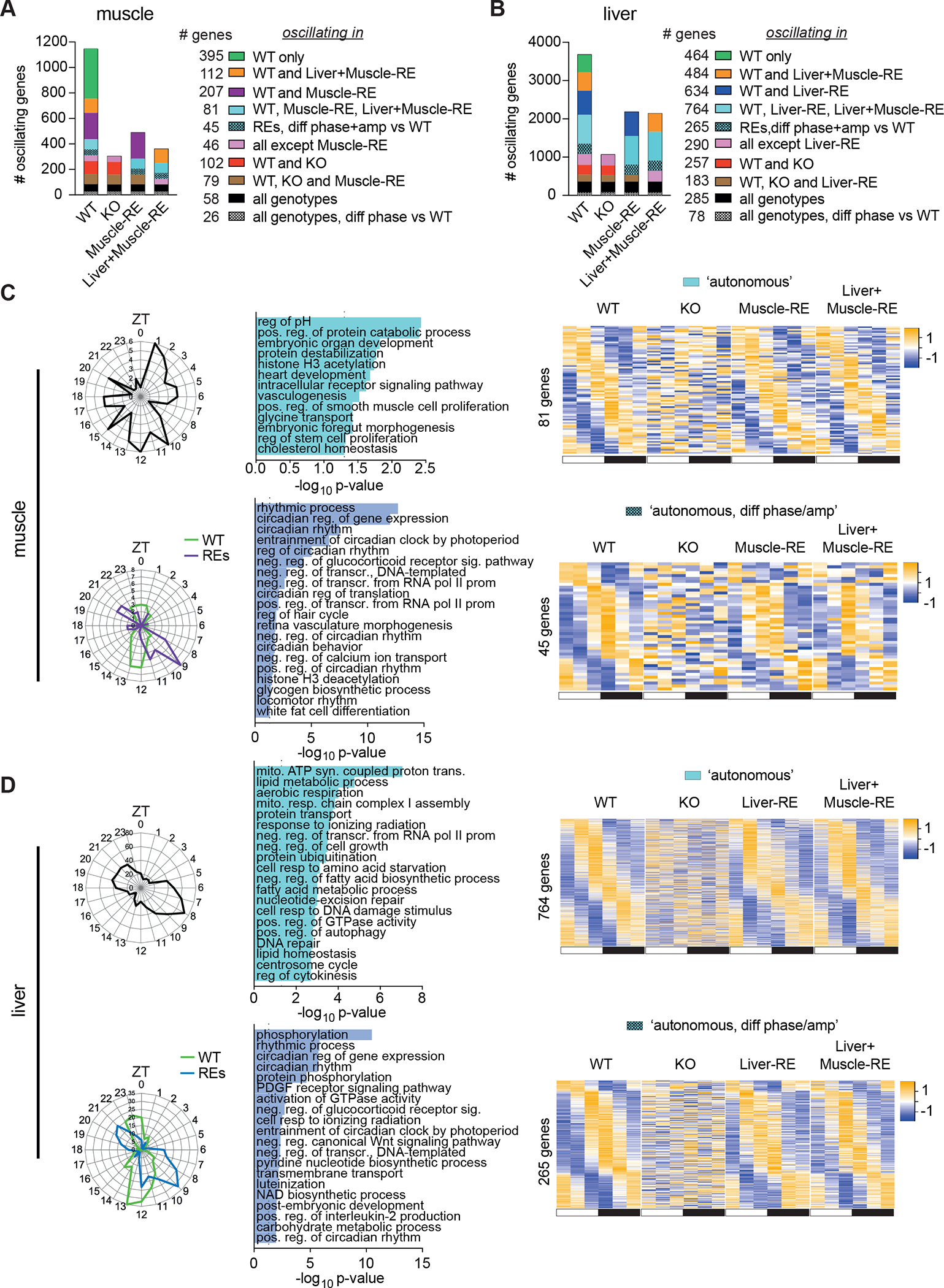
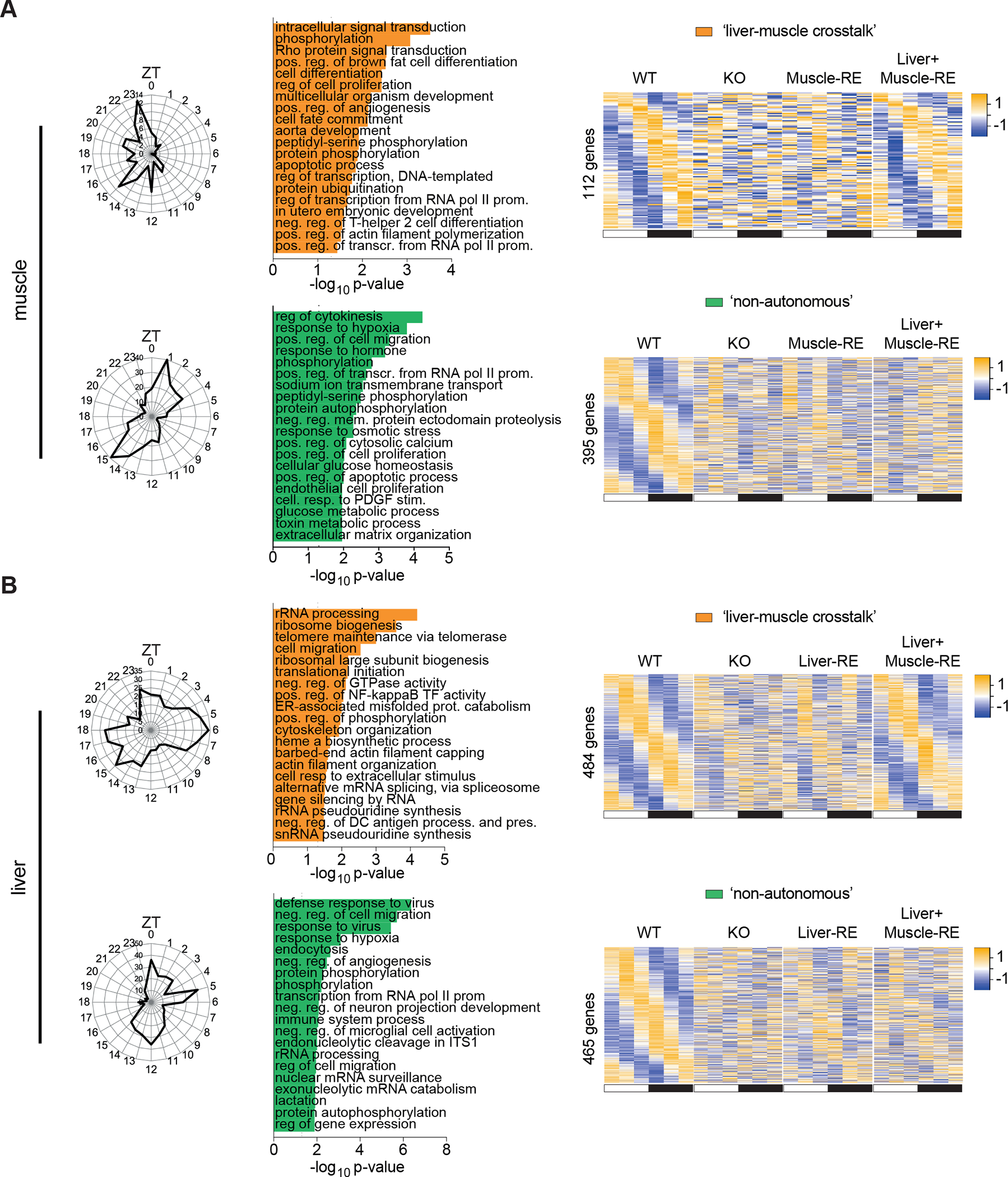
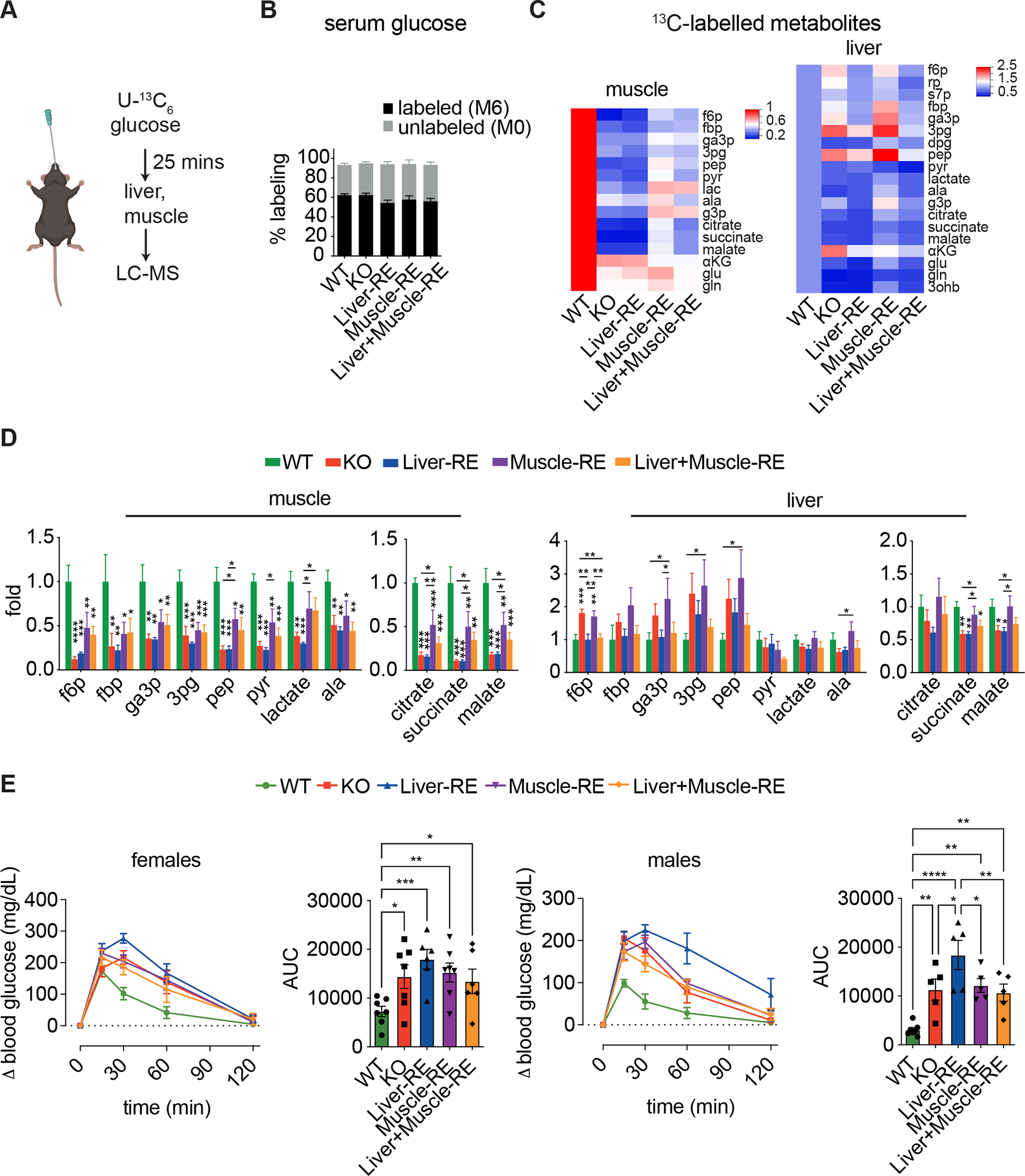
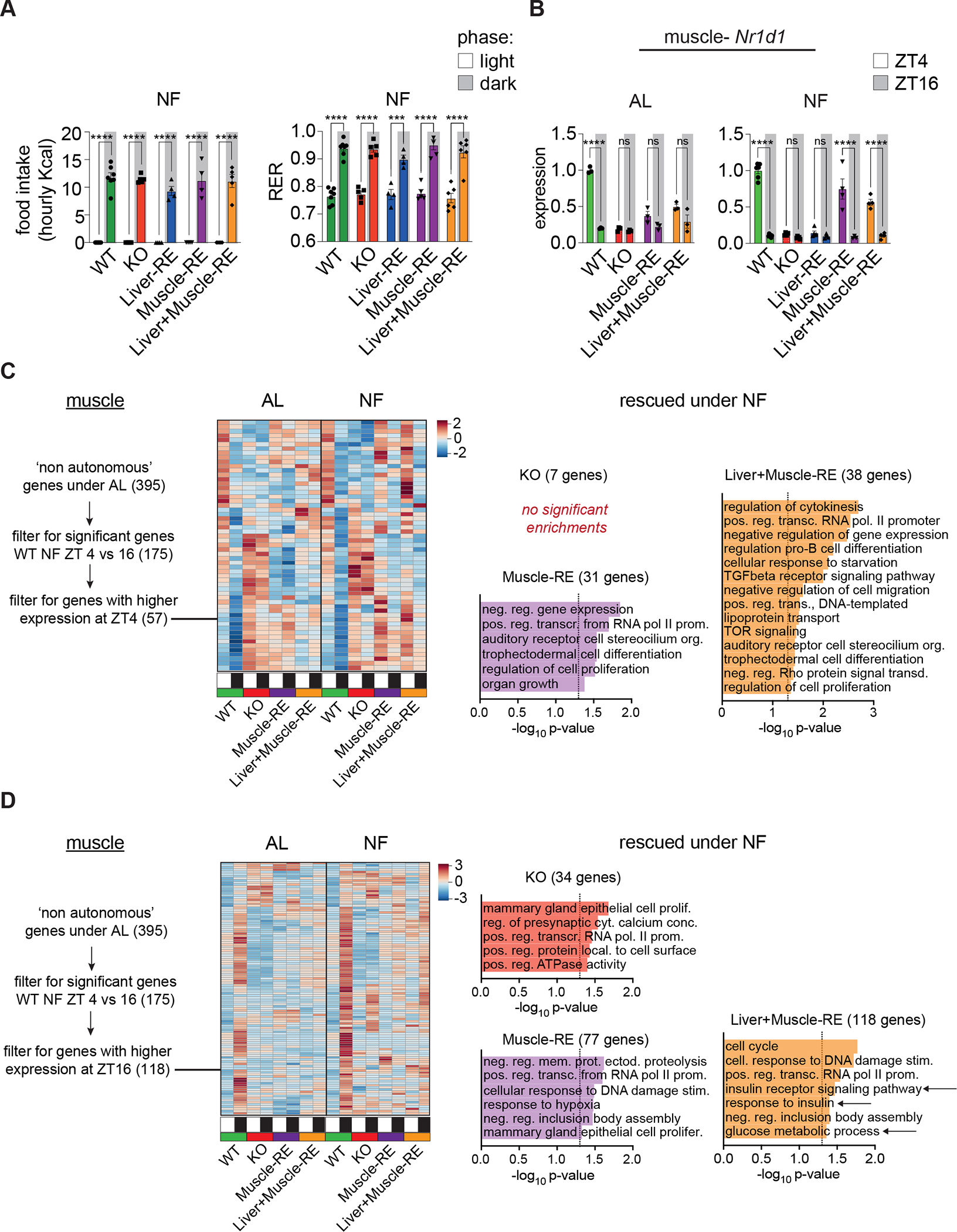
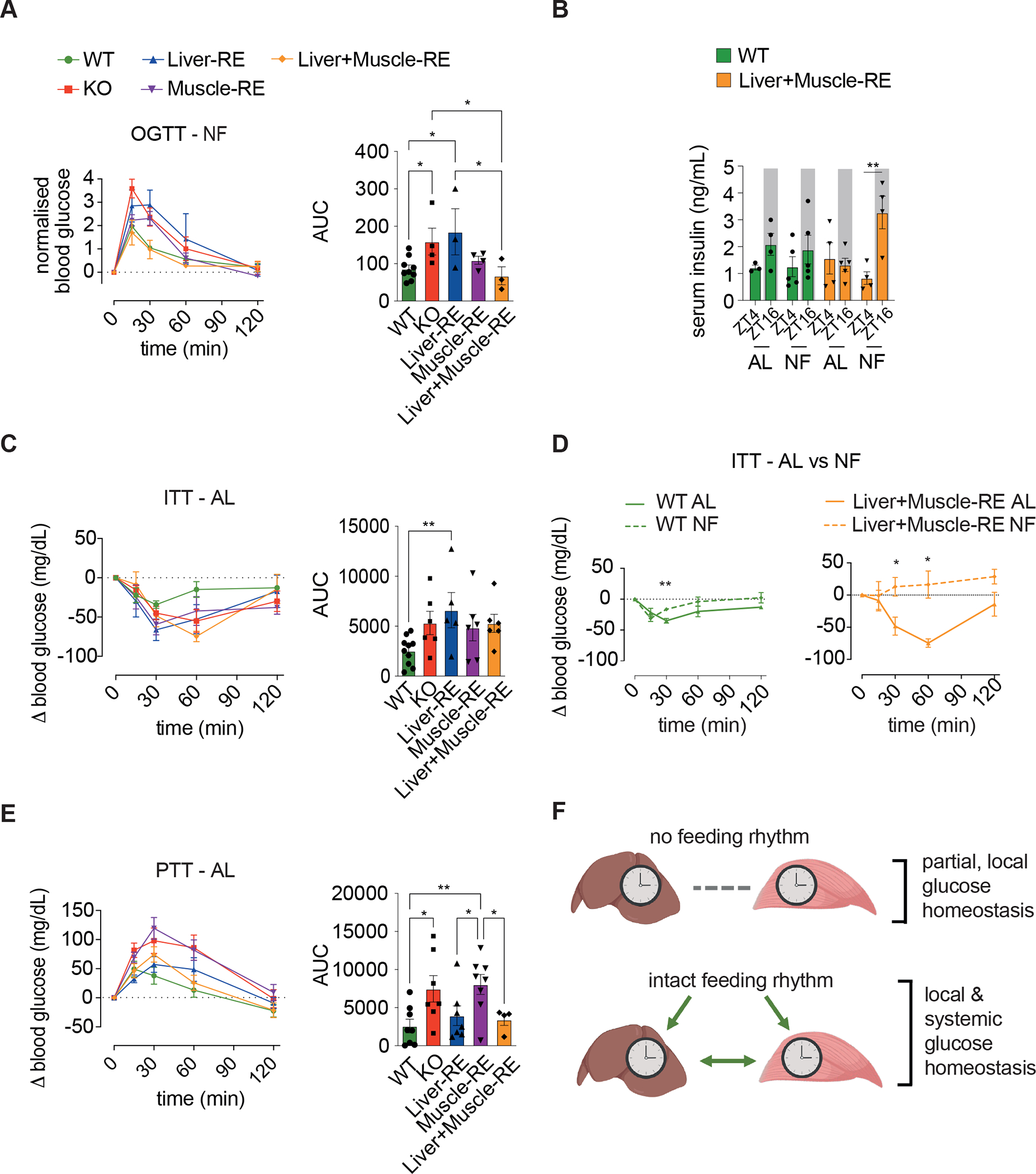
Physiology is regulated by interconnected cell and tissue circadian clocks. Disruption of the rhythms generated by the concerted activity of these clocks is associated with metabolic disease. Here we tested the interactions between clocks in two critical components of organismal metabolism, liver and skeletal muscle, by rescuing clock function either in each organ separately or in both organs simultaneously in otherwise clock-less mice. Experiments showed that individual clocks are partially sufficient for tissue glucose metabolism, yet the connections between both tissue clocks coupled to daily feeding rhythms support systemic glucose tolerance. This synergy relies in part on local transcriptional control of the glucose machinery, feeding-responsive signals such as insulin, and metabolic cycles that connect the muscle and liver. We posit that spatiotemporal mechanisms of muscle and liver play an essential role in the maintenance of systemic glucose homeostasis and that disrupting this diurnal coordination can contribute to metabolic disease.
Keywords: Bmal1; CP: Metabolism; autonomy; circadian rhythms; endocrinology; glucose; inter-organ crosstalk; liver; metabolism; muscle; systems biology.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.A.B. is a co-founder and scientific advisor of ONA Therapeutics.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA244519/CA/NCI NIH HHS/United States
- S10 OD021718/OD/NIH HHS/United States
- R35 GM150618/GM/NIGMS NIH HHS/United States
- T32 CA009054/CA/NCI NIH HHS/United States
- P30 AG013319/AG/NIA NIH HHS/United States
- S10 RR025496/RR/NCRR NIH HHS/United States
- K22 CA212045/CA/NCI NIH HHS/United States
- R21 DK114652/DK/NIDDK NIH HHS/United States
- S10 OD010794/OD/NIH HHS/United States
- P30 CA062203/CA/NCI NIH HHS/United States
- R01 CA259370/CA/NCI NIH HHS/United States
- R01 GM123558/GM/NIGMS NIH HHS/United States
- R01 AA029124/AA/NIAAA NIH HHS/United States
- F32 DK121425/DK/NIDDK NIH HHS/United States
- R21 AG053592/AG/NIA NIH HHS/United States
- T32 NS045540/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
