

Thioredoxin TRXo1 is involved in ABA perception via PYR1 redox regulation
- PMID: 37269685
- PMCID: PMC10244696
- DOI: 10.1016/j.redox.2023.102750
Thioredoxin TRXo1 is involved in ABA perception via PYR1 redox regulation
Abstract
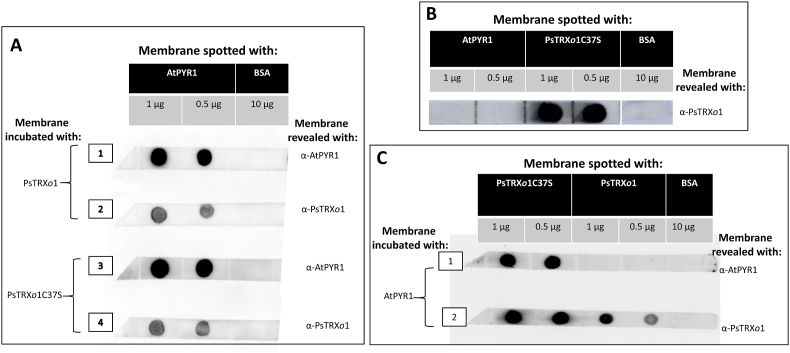
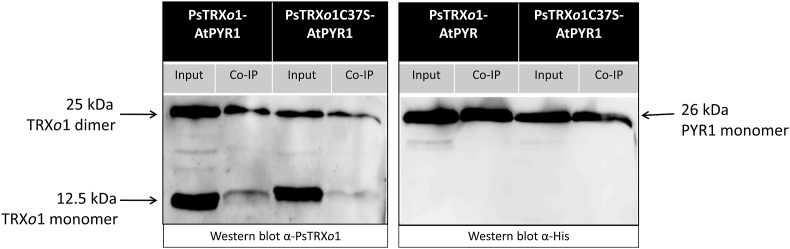
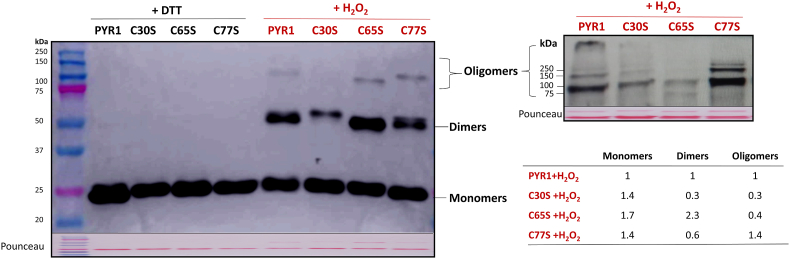
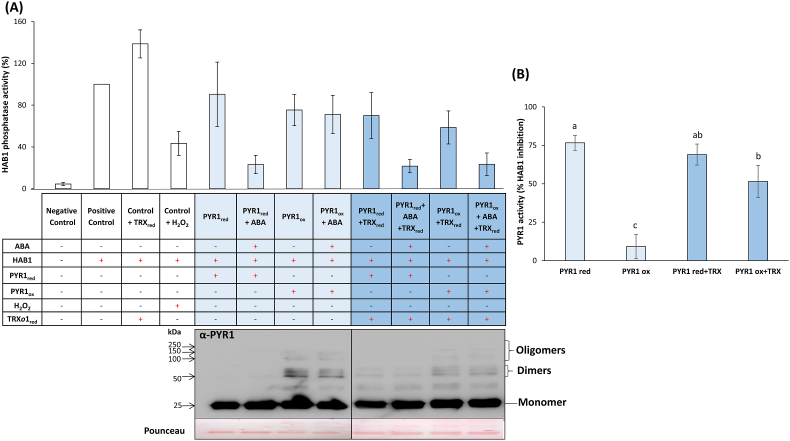
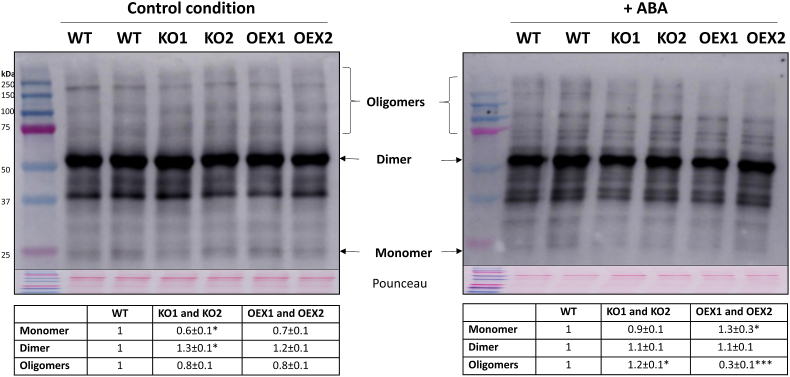
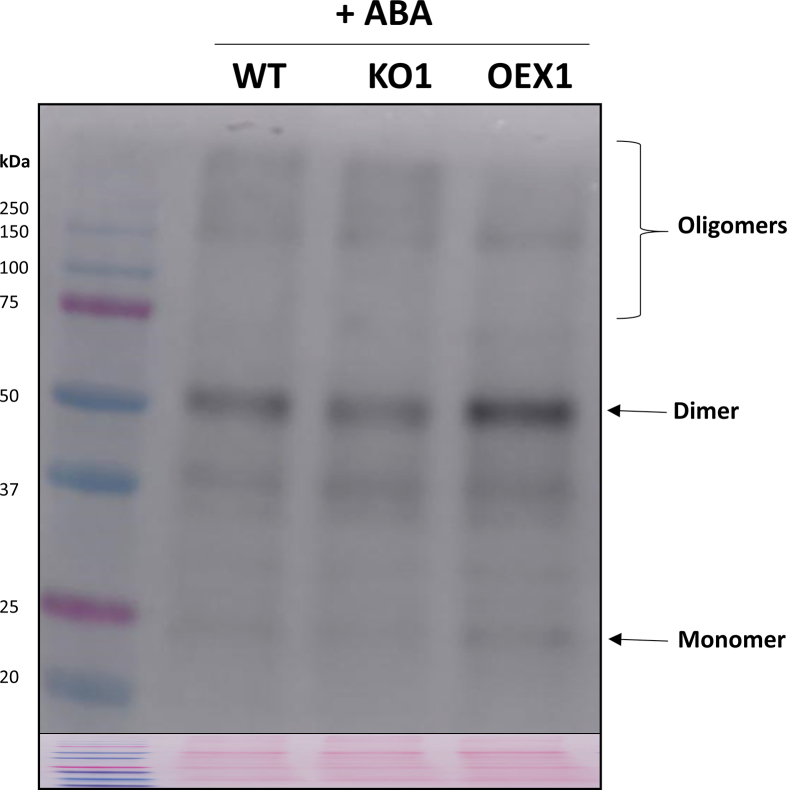
Abscisic acid (ABA) plays a fundamental role in plant growth and development processes such as seed germination, stomatal response or adaptation to stress, amongst others. Increases in the endogenous ABA content is recognized by specific receptors of the PYR/PYL/RCAR family that are coupled to a phosphorylation cascade targeting transcription factors and ion channels. Just like other receptors of the family, nuclear receptor PYR1 binds ABA and inhibits the activity of type 2C phosphatases (PP2Cs), thus avoiding the phosphatase-exerted inhibition on SnRK2 kinases, positive regulators which phosphorylate targets and trigger ABA signalling. Thioredoxins (TRXs) are key components of cellular redox homeostasis that regulate specific target proteins through a thiol-disulfide exchange, playing an essential role in redox homeostasis, cell survival, and growth. In higher plants, TRXs have been found in almost all cellular compartments, although its presence and role in nucleus has been less studied. In this work, affinity chromatography, Dot-blot, co-immunoprecipitation, and bimolecular fluorescence complementation assays allowed us to identify PYR1 as a new TRXo1 target in the nucleus. Studies on recombinant HisAtPYR1 oxidation-reduction with wild type and site-specific mutagenized forms showed that the receptor underwent redox regulation involving changes in the oligomeric state in which Cys30 and Cys65 residues were implied. TRXo1 was able to reduce previously-oxidized inactive PYR1, thus recovering its capacity to inhibit HAB1 phosphatase. In vivo PYR1 oligomerization was dependent on the redox state, and a differential pattern was detected in KO and over-expressing Attrxo1 mutant plants grown in the presence of ABA compared to WT plants. Thus, our findings suggest the existence of a redox regulation of TRXo1 on PYR1 that may be relevant for ABA signalling and had not been described so far.
Keywords: Abscisic acid; Oligomerization; PYR1; Protein interaction; Redox regulation; Thioredoxin o1.
Copyright © 2023 Consejo Superior de Investigaciones Científicas. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest None.
Figures










Similar articles
-
PYR/PYL/RCAR family members are major in-vivo ABI1 protein phosphatase 2C-interacting proteins in Arabidopsis.Plant J. 2010 Jan;61(2):290-9. doi: 10.1111/j.1365-313X.2009.04054.x. Epub 2009 Oct 26. Plant J. 2010. PMID: 19874541 Free PMC article.
-
Arabidopsis PYR/PYL/RCAR receptors play a major role in quantitative regulation of stomatal aperture and transcriptional response to abscisic acid.Plant Cell. 2012 Jun;24(6):2483-96. doi: 10.1105/tpc.112.098574. Epub 2012 Jun 26. Plant Cell. 2012. PMID: 22739828 Free PMC article.
-
Modulation of abscisic acid signaling in vivo by an engineered receptor-insensitive protein phosphatase type 2C allele.Plant Physiol. 2011 May;156(1):106-16. doi: 10.1104/pp.110.170894. Epub 2011 Feb 28. Plant Physiol. 2011. PMID: 21357183 Free PMC article.
-
Structural insights into PYR/PYL/RCAR ABA receptors and PP2Cs.Plant Sci. 2012 Jan;182:3-11. doi: 10.1016/j.plantsci.2010.11.014. Epub 2010 Dec 7. Plant Sci. 2012. PMID: 22118610 Review.
-
Omics Approaches Toward Defining the Comprehensive Abscisic Acid Signaling Network in Plants.Plant Cell Physiol. 2015 Jun;56(6):1043-52. doi: 10.1093/pcp/pcv060. Epub 2015 Apr 26. Plant Cell Physiol. 2015. PMID: 25917608 Review.
Cited by
-
Reactive Byproducts of Plant Redox Metabolism and Protein Functions.Acta Naturae. 2024 Oct-Dec;16(4):48-61. doi: 10.32607/actanaturae.27477. Acta Naturae. 2024. PMID: 39877007 Free PMC article.
-
A fungal pathogen suppresses host leaf senescence to increase infection.Nat Commun. 2025 Mar 24;16(1):2864. doi: 10.1038/s41467-025-58277-5. Nat Commun. 2025. PMID: 40128252 Free PMC article.
-
New role for thioredoxins in plants: Implication of TRXo1 in protein depersulfidation.Redox Biol. 2025 May;82:103627. doi: 10.1016/j.redox.2025.103627. Epub 2025 Apr 7. Redox Biol. 2025. PMID: 40220625 Free PMC article.
-
Redox regulation, thioredoxins, and glutaredoxins in retrograde signalling and gene transcription.J Exp Bot. 2023 Oct 13;74(19):5955-5969. doi: 10.1093/jxb/erad270. J Exp Bot. 2023. PMID: 37453076 Free PMC article. Review.
-
Combining genotyping approaches improves resolution for association mapping: a case study in tropical maize under water stress conditions.Front Plant Sci. 2025 Jan 23;15:1442008. doi: 10.3389/fpls.2024.1442008. eCollection 2024. Front Plant Sci. 2025. PMID: 39917602 Free PMC article.
References
-
- Sakata Y., Komatsu K., Takezawa D. In: Progress in Botany 75. Lüttge U., Beyschlag W., Cushman J., editors. Springer-Verlag, Berlin Heidelberg; 2014. ABA as a universal plant hormone; pp. 57–96. 2014. - DOI
-
- Ruggiero A., Landi S., Punzo P., Possenti M., Van Oosten M.J., Costa A., Morelli G., Maggio A., Grillo S., Batelli G. Salinity and ABA seed responses in pepper: expression and interaction of ABA core signaling components. Front. Plant Sci. 2019;10:304. doi: 10.3389/fpls.2019.00304. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
