

The role of microglial LRRK2 kinase in manganese-induced inflammatory neurotoxicity via NLRP3 inflammasome and RAB10-mediated autophagy dysfunction
- PMID: 37269951
- PMCID: PMC10331485
- DOI: 10.1016/j.jbc.2023.104879
The role of microglial LRRK2 kinase in manganese-induced inflammatory neurotoxicity via NLRP3 inflammasome and RAB10-mediated autophagy dysfunction
Abstract
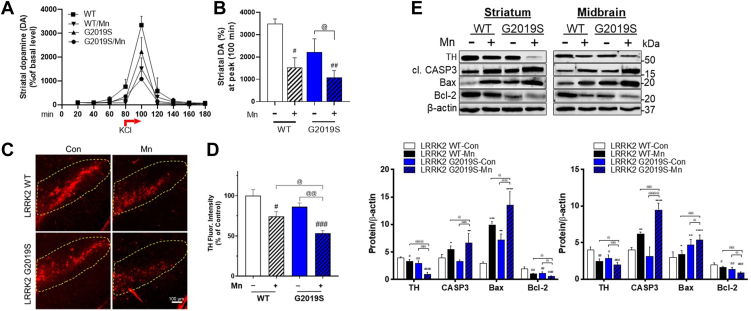
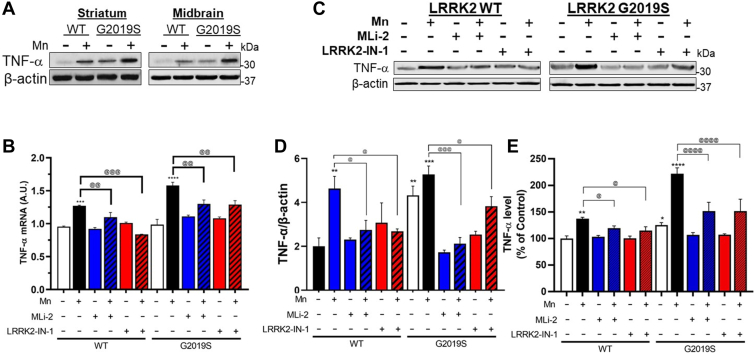
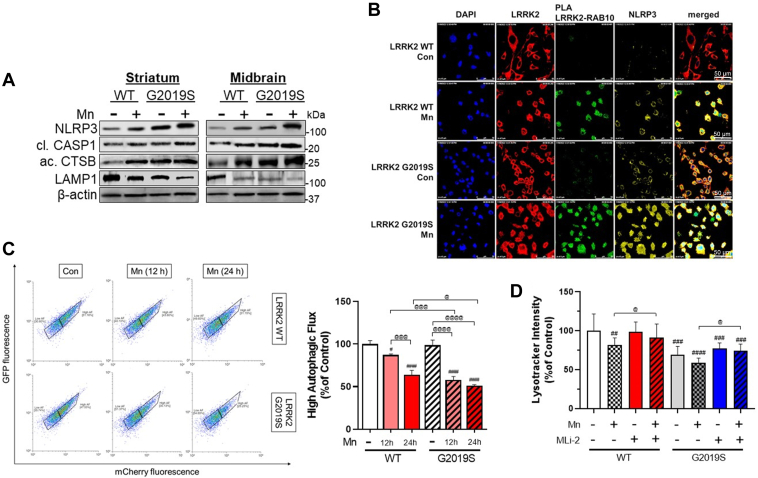
Chronic manganese (Mn) exposure can lead to manganism, a neurological disorder sharing common symptoms with Parkinson's disease (PD). Studies have shown that Mn can increase the expression and activity of leucine-rich repeat kinase 2 (LRRK2), leading to inflammation and toxicity in microglia. LRRK2 G2019S mutation also elevates LRRK2 kinase activity. Thus, we tested if Mn-increased microglial LRRK2 kinase is responsible for Mn-induced toxicity, and exacerbated by G2019S mutation, using WT and LRRK2 G2019S knock-in mice and BV2 microglia. Mn (30 mg/kg, nostril instillation, daily for 3 weeks) caused motor deficits, cognitive impairments, and dopaminergic dysfunction in WT mice, which were exacerbated in G2019S mice. Mn induced proapoptotic Bax, NLRP3 inflammasome, IL-1β, and TNF-α in the striatum and midbrain of WT mice, and these effects were more pronounced in G2019S mice. BV2 microglia were transfected with human LRRK2 WT or G2019S, followed by Mn (250 μM) exposure to better characterize its mechanistic action. Mn increased TNF-α, IL-1β, and NLRP3 inflammasome activation in BV2 cells expressing WT LRRK2, which was elevated further in G2019S-expressing cells, while pharmacological inhibition of LRRK2 mitigated these effects in both genotypes. Moreover, the media from Mn-treated G2019S-expressing BV2 microglia caused greater toxicity to the cath.a-differentiated (CAD) neuronal cells compared to media from microglia expressing WT. Mn-LRRK2 activated RAB10 which was exacerbated in G2019S. RAB10 played a critical role in LRRK2-mediated Mn toxicity by dysregulating the autophagy-lysosome pathway and NLRP3 inflammasome in microglia. Our novel findings suggest that microglial LRRK2 via RAB10 plays a critical role in Mn-induced neuroinflammation.
Keywords: G2019S; LRRK2; NLRP3 inflammasome; RAB10; autophagy; inflammation; lysosome; manganese; microglia.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare no conflicts of interest with the contents of this article.
Figures









Update of
-
The role of microglial LRRK2 in manganese-induced inflammatory neurotoxicity via NLRP3 inflammasome and RAB10-mediated autophagy dysfunction.bioRxiv [Preprint]. 2023 Apr 5:2023.04.03.535418. doi: 10.1101/2023.04.03.535418. bioRxiv. 2023. Update in: J Biol Chem. 2023 Jul;299(7):104879. doi: 10.1016/j.jbc.2023.104879. PMID: 37066140 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
