

The composition of environmental microbiota in three tree fruit packing facilities changed over seasons and contained taxa indicative of L. monocytogenes contamination
- PMID: 37271802
- PMCID: PMC10240739
- DOI: 10.1186/s40168-023-01544-8
The composition of environmental microbiota in three tree fruit packing facilities changed over seasons and contained taxa indicative of L. monocytogenes contamination
Abstract
Background: Listeria monocytogenes can survive in cold and wet environments, such as tree fruit packing facilities and it has been implicated in outbreaks and recalls of tree fruit products. However, little is known about microbiota that co-occurs with L. monocytogenes and its stability over seasons in tree fruit packing environments. In this 2-year longitudinal study, we aimed to characterize spatial and seasonal changes in microbiota composition and identify taxa indicative of L. monocytogenes contamination in wet processing areas of three tree fruit packing facilities (F1, F2, F3).
Methods: A total of 189 samples were collected during two apple packing seasons from floors under the washing, drying, and waxing areas. The presence of L. monocytogenes was determined using a standard culturing method, and environmental microbiota was characterized using amplicon sequencing. PERMANOVA was used to compare microbiota composition among facilities over two seasons, and abundance-occupancy analysis was used to identify shared and temporal core microbiota. Differential abundance analysis and random forest were applied to detect taxa indicative of L. monocytogenes contamination. Lastly, three L. monocytogenes-positive samples were sequenced using shotgun metagenomics with Nanopore MinION, as a proof-of-concept for direct detection of L. monocytogenes' DNA in environmental samples.
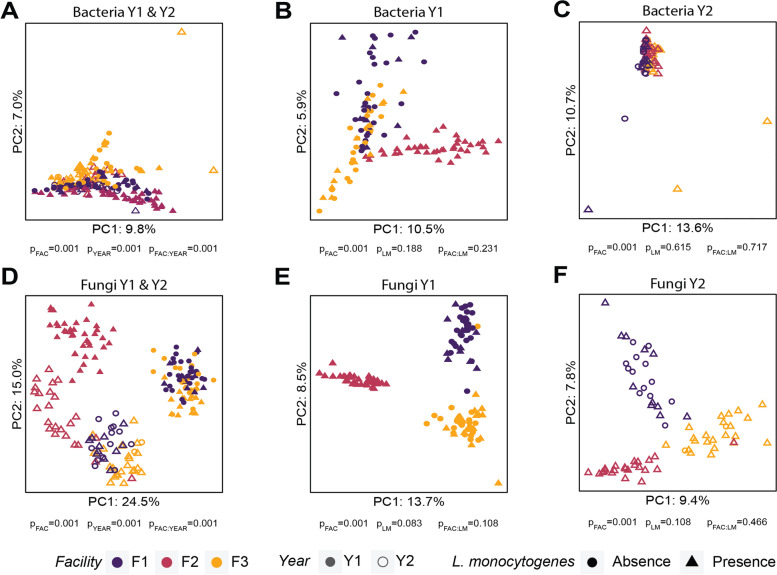
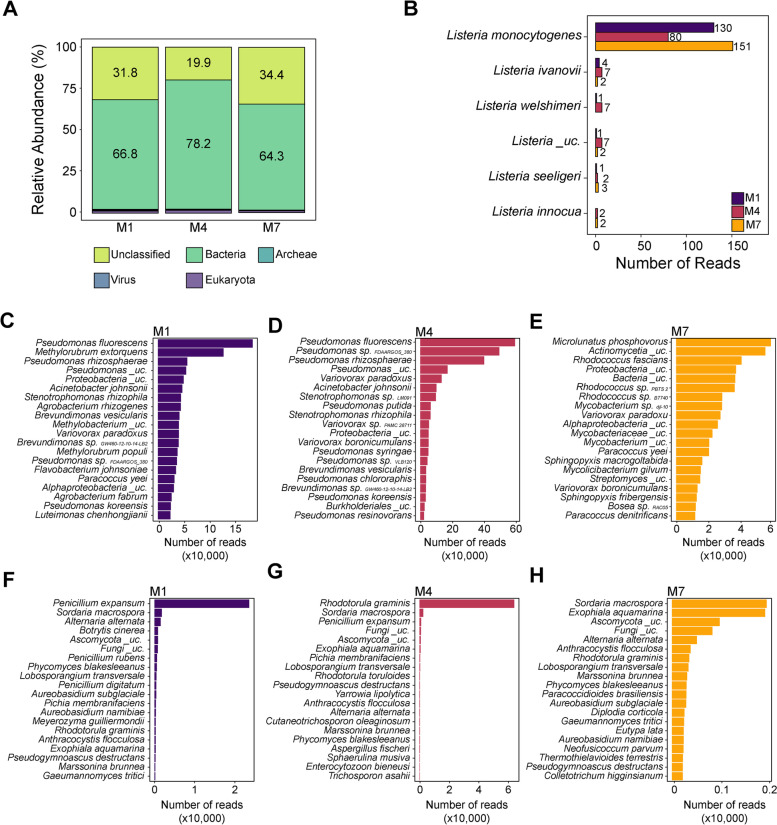
Results: The occurrence of L. monocytogenes significantly increased from 28% in year 1 to 46% in year 2 in F1, and from 41% in year 1 to 92% in year 2 in F3, while all samples collected from F2 were L. monocytogenes-positive in both years. Samples collected from three facilities had a significantly different microbiota composition in both years, but the composition of each facility changed over years. A subset of bacterial taxa including Pseudomonas, Stenotrophomonas, and Microbacterium, and fungal taxa, including Yarrowia, Kurtzmaniella, Cystobasidium, Paraphoma, and Cutaneotrichosporon, were identified as potential indicators of L. monocytogenes within the monitored environments. Lastly, the DNA of L. monocytogenes was detected through direct Nanopore sequencing of metagenomic DNA extracted from environmental samples.
Conclusions: This study demonstrated that a cross-sectional sampling strategy may not accurately reflect the representative microbiota of food processing facilities. Our findings also suggest that specific microorganisms are indicative of L. monocytogenes, warranting further investigation of their role in the survival and persistence of L. monocytogenes. Video Abstract.
Keywords: Food safety; Listeria monocytogenes; Microbiota; Spatial and temporal variation; Tree fruit packing facilities.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures





References
-
- Angelo KM, Conrad AR, Saupe A, Dragoo H, West N, Sorenson A, et al. Multistate outbreak of Listeria monocytogenes infections linked to whole apples used in commercially produced, prepackaged caramel apples: United States, 2014–2015. Epidemiol Infect. 2017;145:848–856. doi: 10.1017/S0950268816003083. - DOI - PMC - PubMed
-
- Gustafson RE, Ryser ET. Thermal inactivation and growth of Listeria monocytogenes during production and storage of caramel apples. Food Control. 2017;79:234–238. doi: 10.1016/j.foodcont.2017.03.043. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
