

Cardiomyopathy related desmocollin-2 prodomain variants affect the intracellular cadherin transport and processing
- PMID: 37273868
- PMCID: PMC10235514
- DOI: 10.3389/fcvm.2023.1127261
Cardiomyopathy related desmocollin-2 prodomain variants affect the intracellular cadherin transport and processing
Abstract
Background: Arrhythmogenic cardiomyopathy can be caused by genetic variants in desmosomal cadherins. Since cardiac desmosomal cadherins are crucial for cell-cell-adhesion, their correct localization at the plasma membrane is essential.
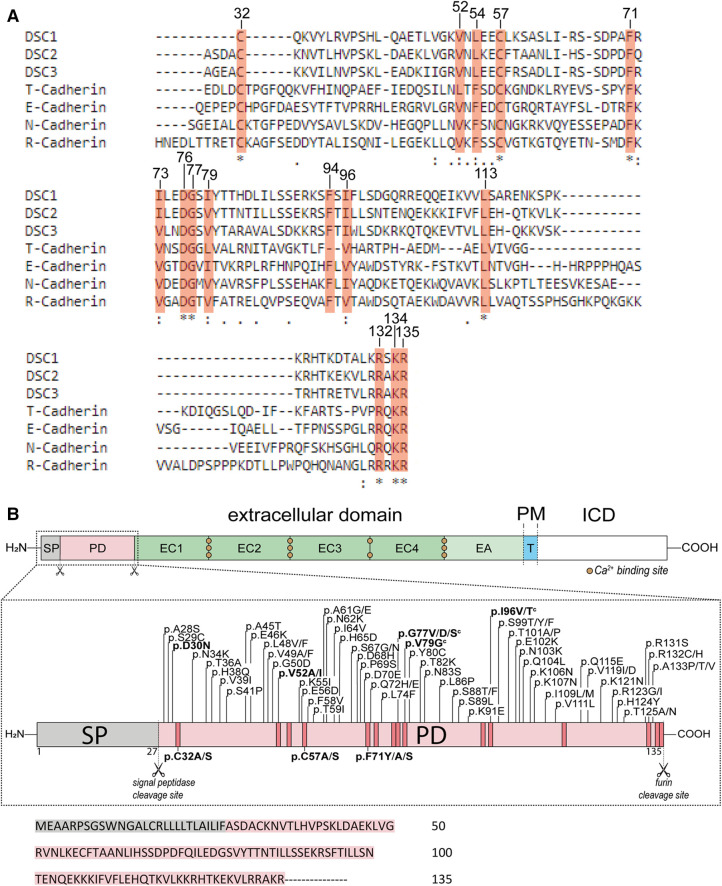
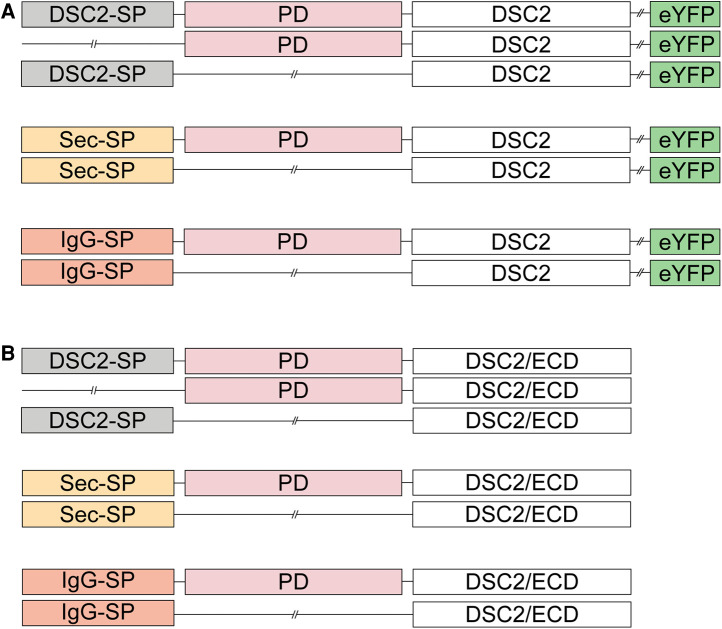
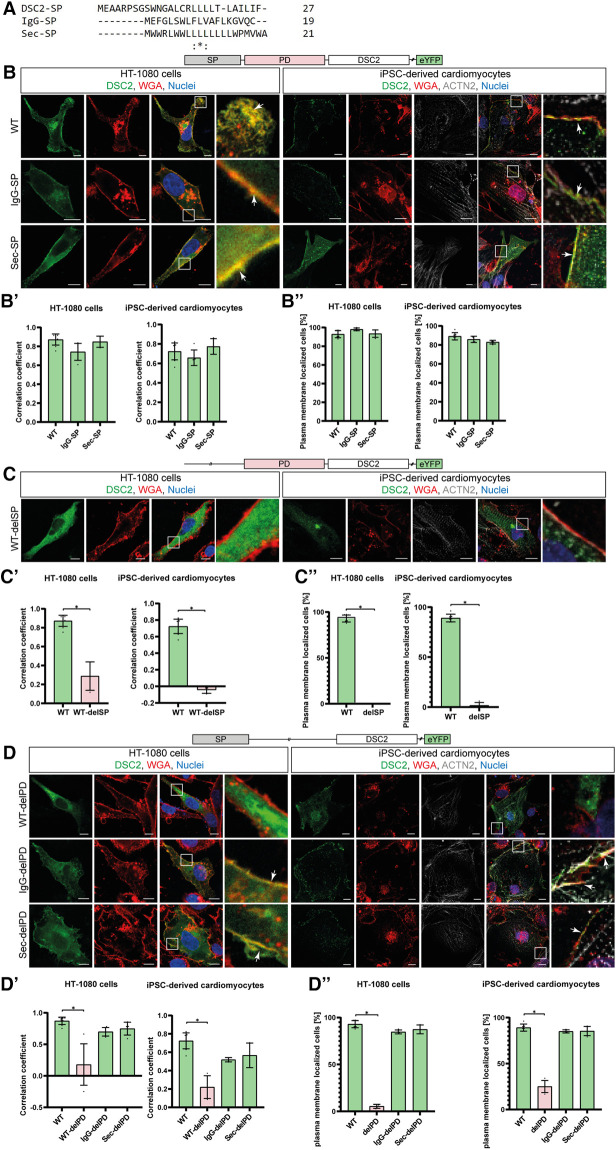
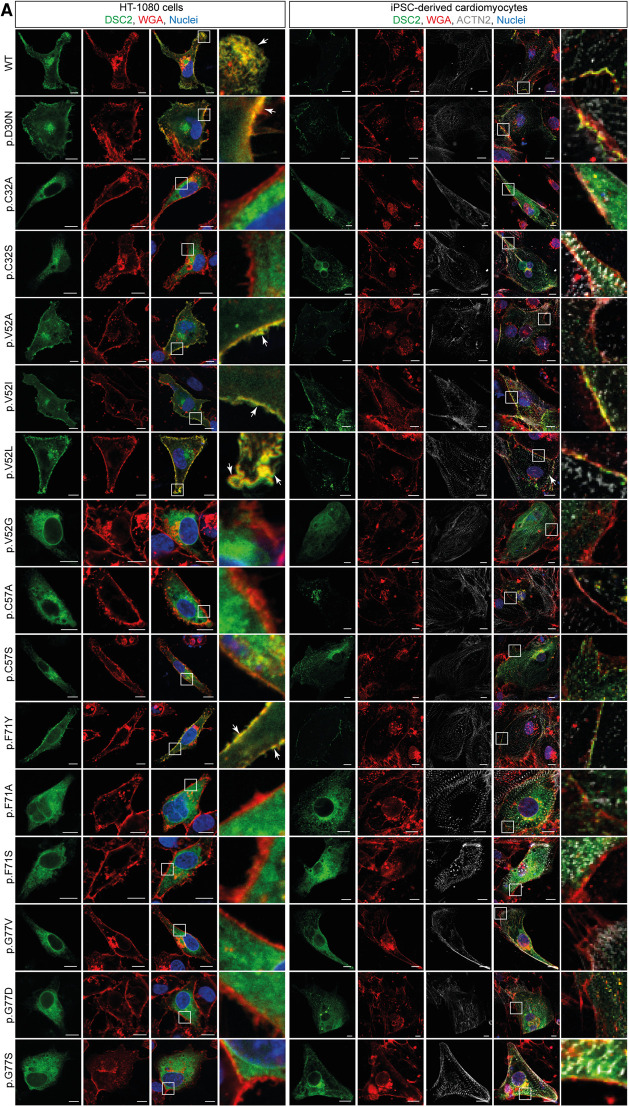
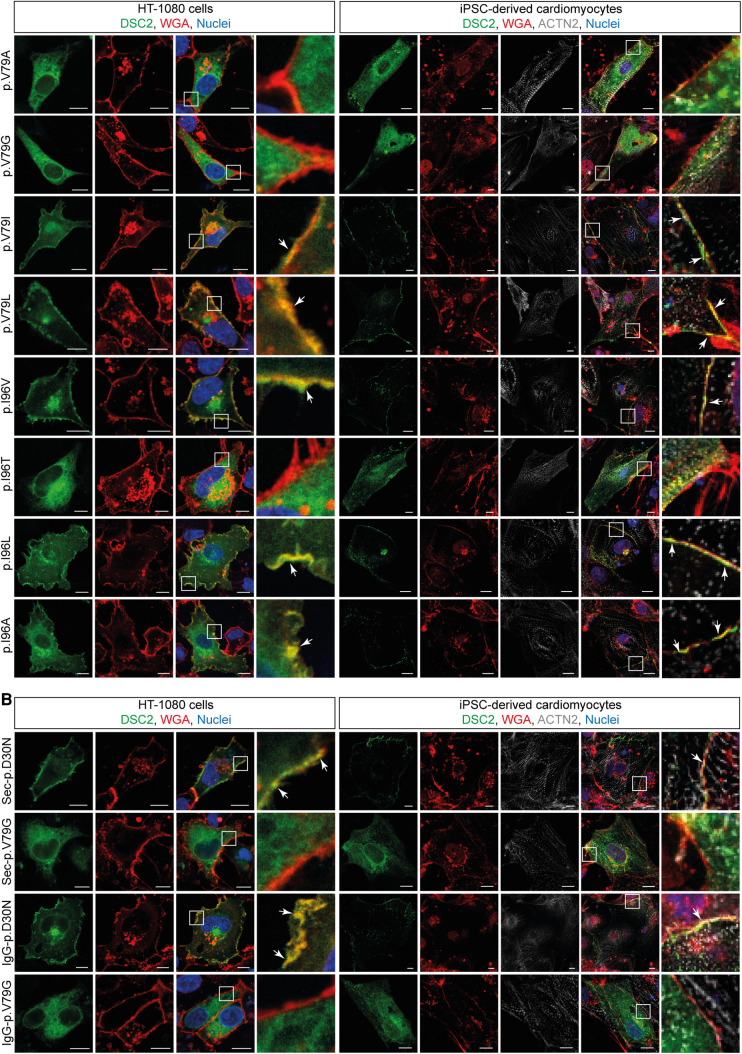
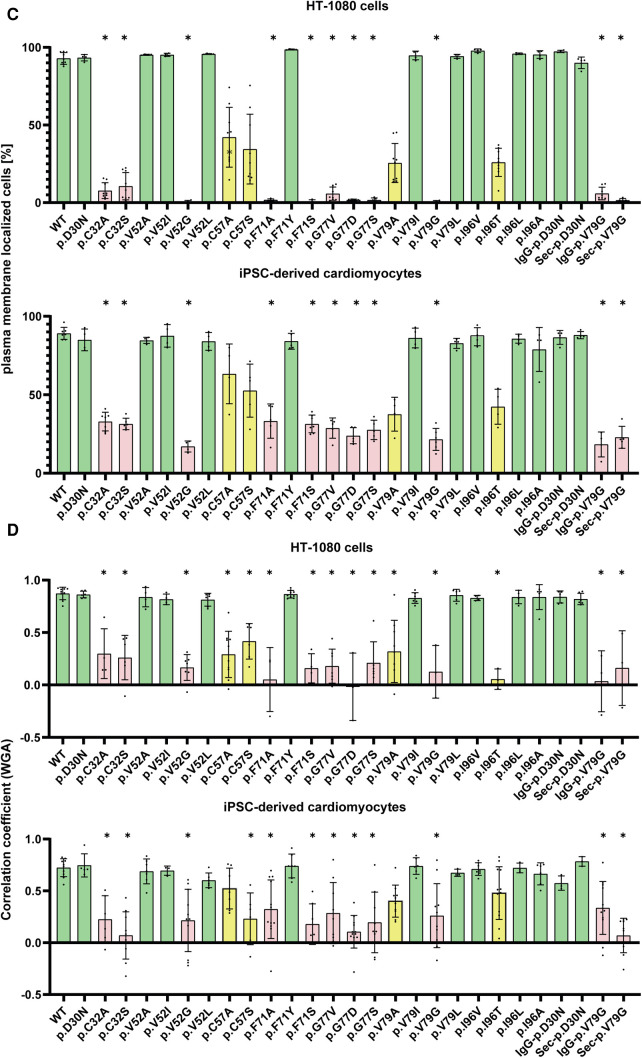
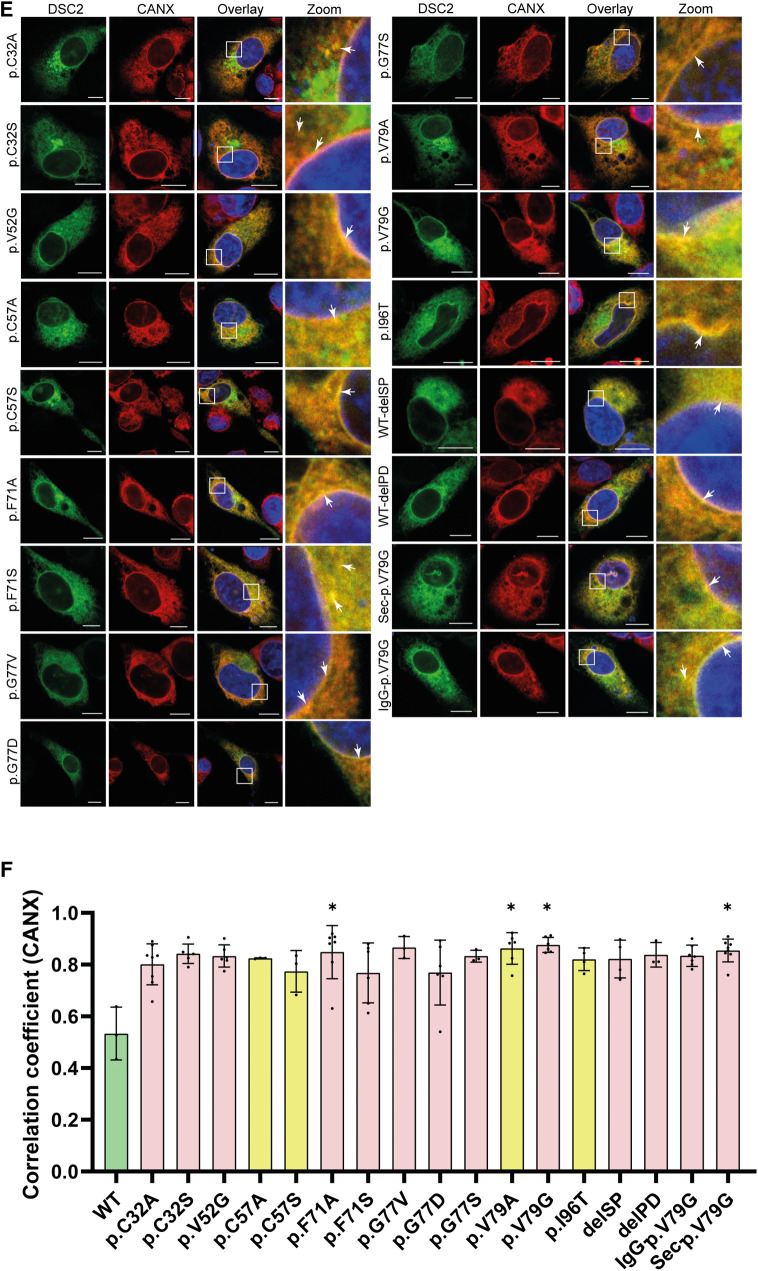
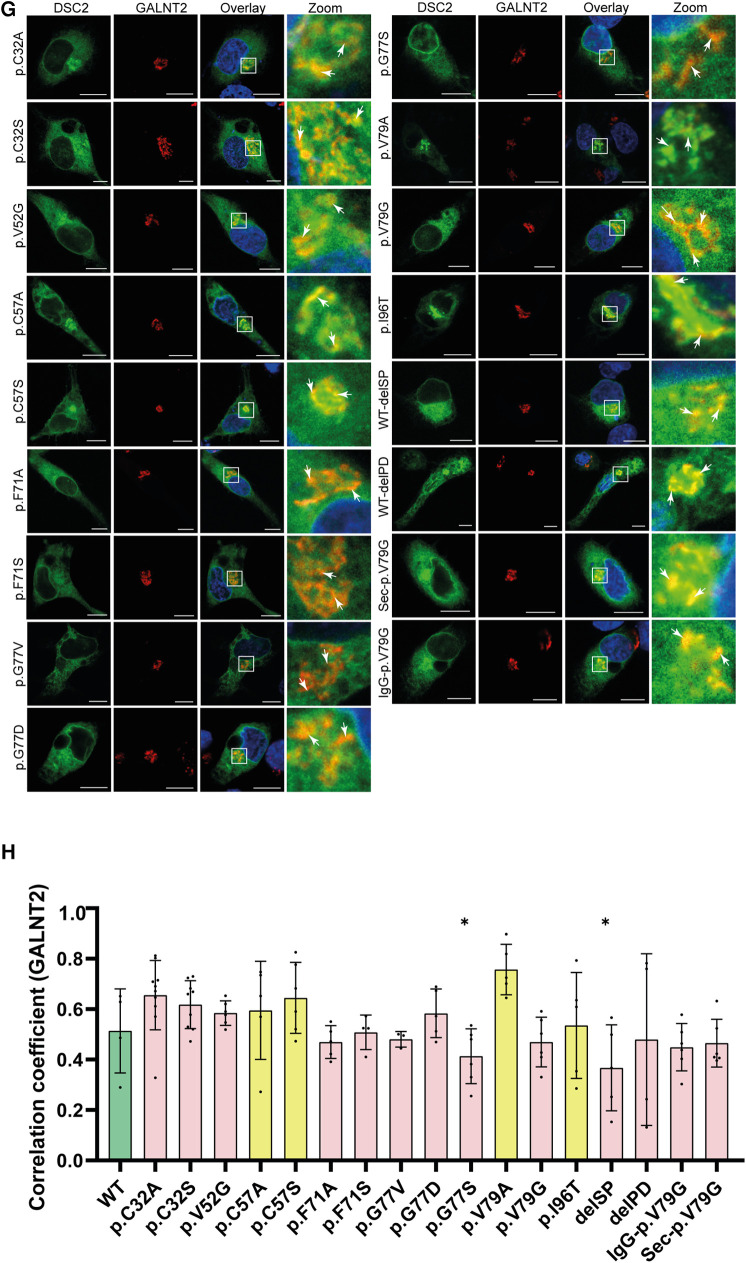
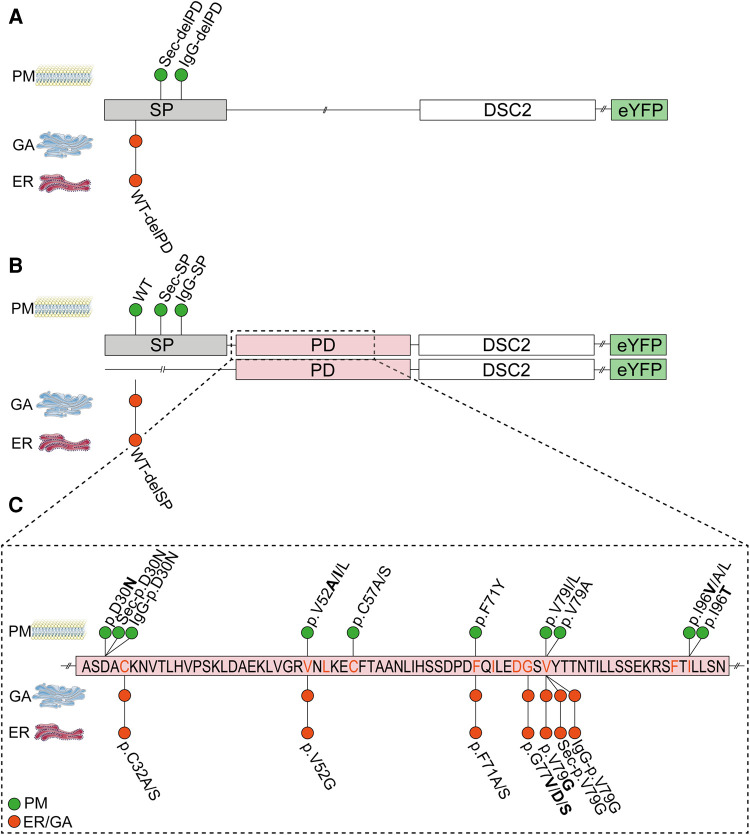
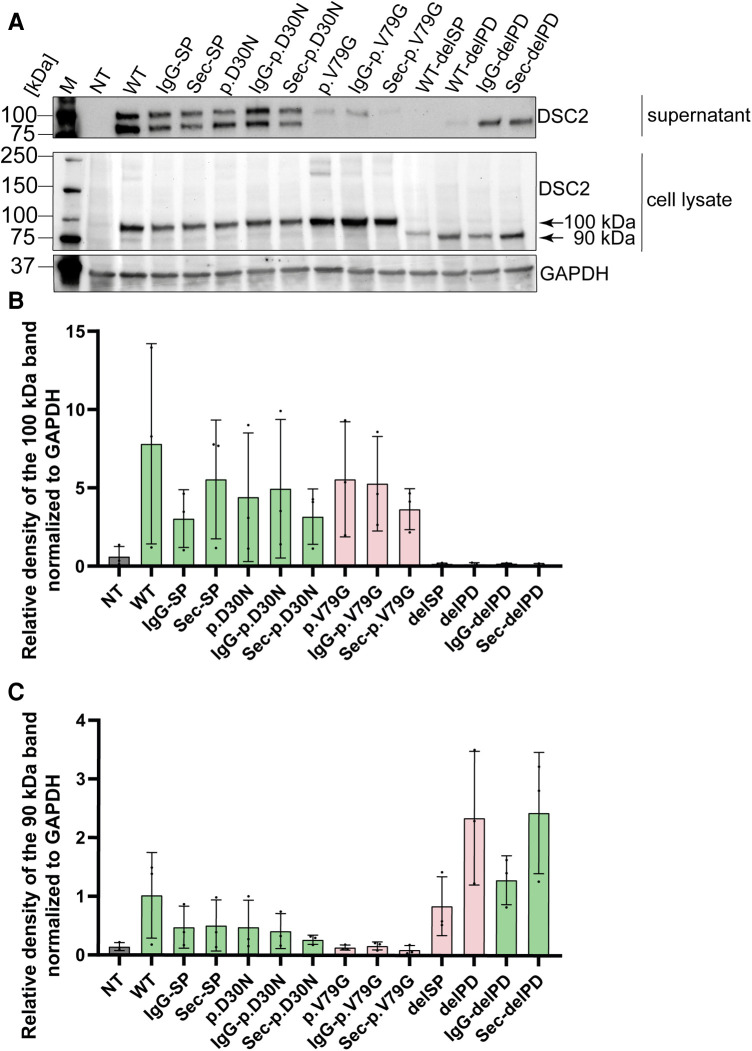
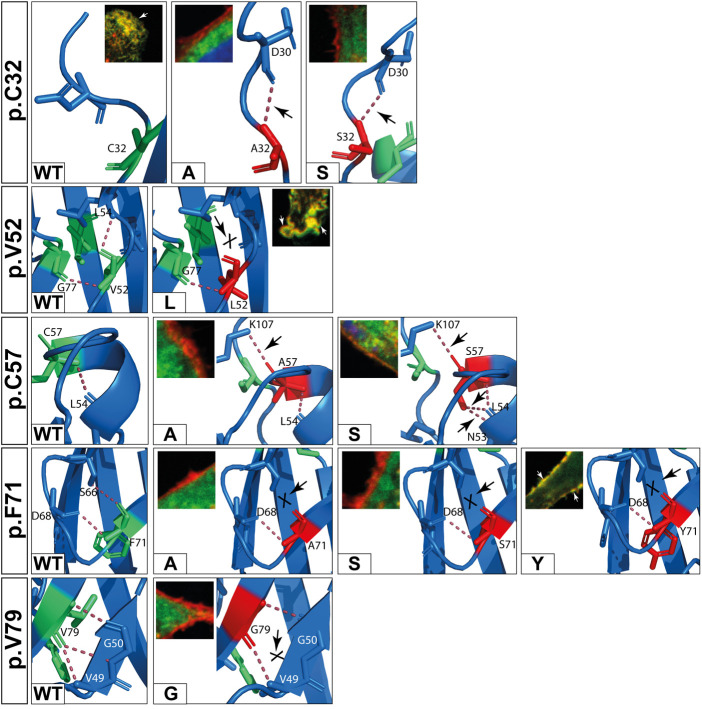
Methods: Nine desmocollin-2 variants at five positions from various public genetic databases (p.D30N, p.V52A/I, p.G77V/D/S, p.V79G, p.I96V/T) and three additional conserved positions (p.C32, p.C57, p.F71) within the prodomain were investigated in vitro using confocal microscopy. Model variants (p.C32A/S, p.V52G/L, p.C57A/S, p.F71Y/A/S, p.V79A/I/L, p.I96l/A) were generated to investigate the impact of specific amino acids.
Results: We revealed that all analyzed positions in the prodomain are critical for the intracellular transport. However, the variants p.D30N, p.V52A/I and p.I96V listed in genetic databases do not disturb the intracellular transport revealing that the loss of these canonical sequences may be compensated.
Conclusion: As disease-related homozygous truncating desmocollin-2 variants lacking the transmembrane domain are not localized at the plasma membrane, we predict that some of the investigated prodomain variants may be relevant in the context of arrhythmogenic cardiomyopathy due to disturbed intracellular transport.
Keywords: DSC2; arrhythmogenic cardiomyopathy; desmocollin-2; desmosome; genetic variants; intracellular transport; prodomain; signal peptide.
© 2023 Pohl, Göz, Gärtner, Brodehl, Cimen, Saguner, Schulze-Bahr, Walhorn, Anselmetti and Milting.
Conflict of interest statement
AMS received educational grants through his institution from Abbott; Bayer Healthcare; Biosense Webster; Biotronik; Boston Scientific; BMS/Pfizer and Medtronic; and speaker/advisory board/consulting fees from Bayer Healthcare; Daiichi-Sankyo; Medtronic; Novartis and Pfizer. AB is a shareholder of Tenaya Therapeutics. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources
Research Materials