

Staphylococcus aureus δ-toxin present on skin promotes the development of food allergy in a murine model
- PMID: 37275864
- PMCID: PMC10235538
- DOI: 10.3389/fimmu.2023.1173069
Staphylococcus aureus δ-toxin present on skin promotes the development of food allergy in a murine model
Abstract
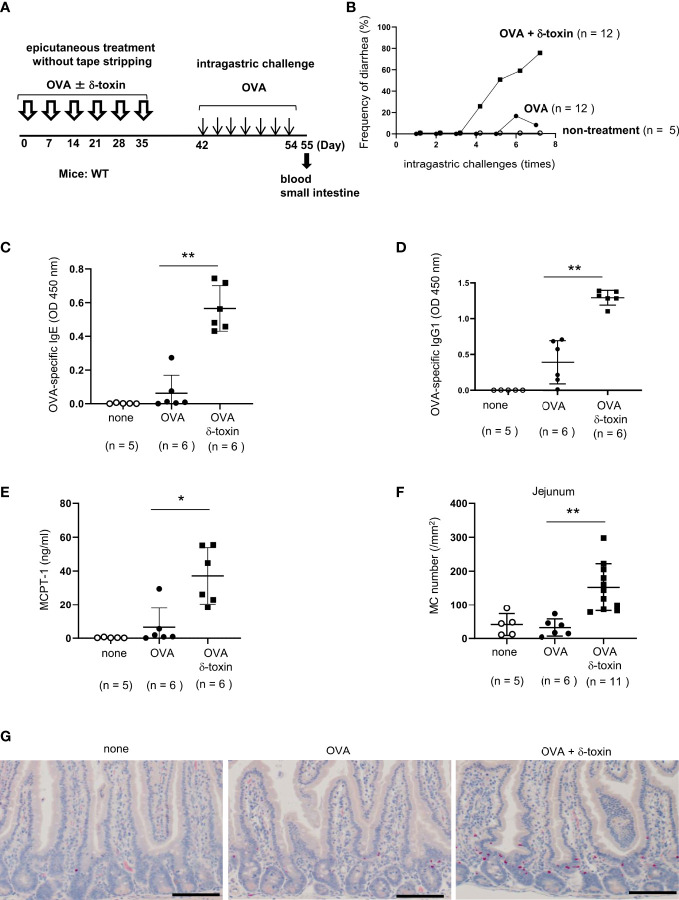
Background: Patients with food allergy often suffer from atopic dermatitis, in which Staphylococcus aureus colonization is frequently observed. Staphylococcus aureus δ-toxin activates mast cells and promotes T helper 2 type skin inflammation in the tape-stripped murine skin. However, the physiological effects of δ-toxin present on the steady-state skin remain unknown. We aimed to investigate whether δ-toxin present on the steady-state skin impacts the development of food allergy.
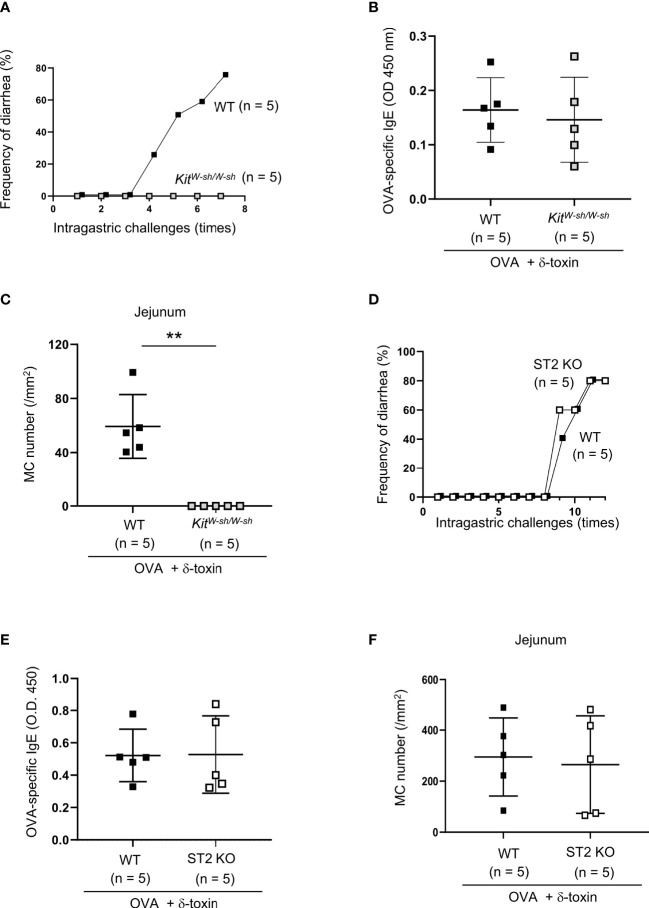
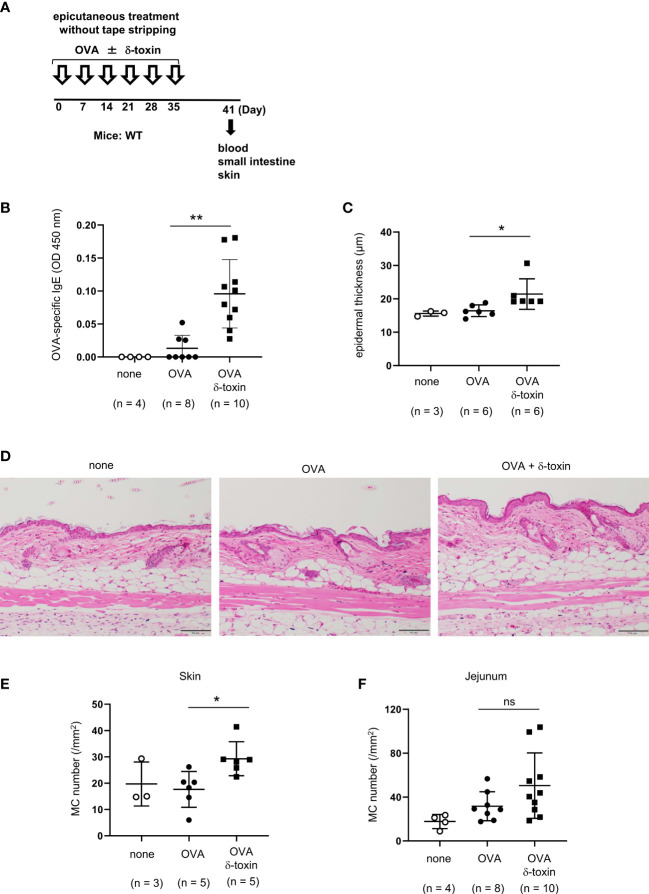
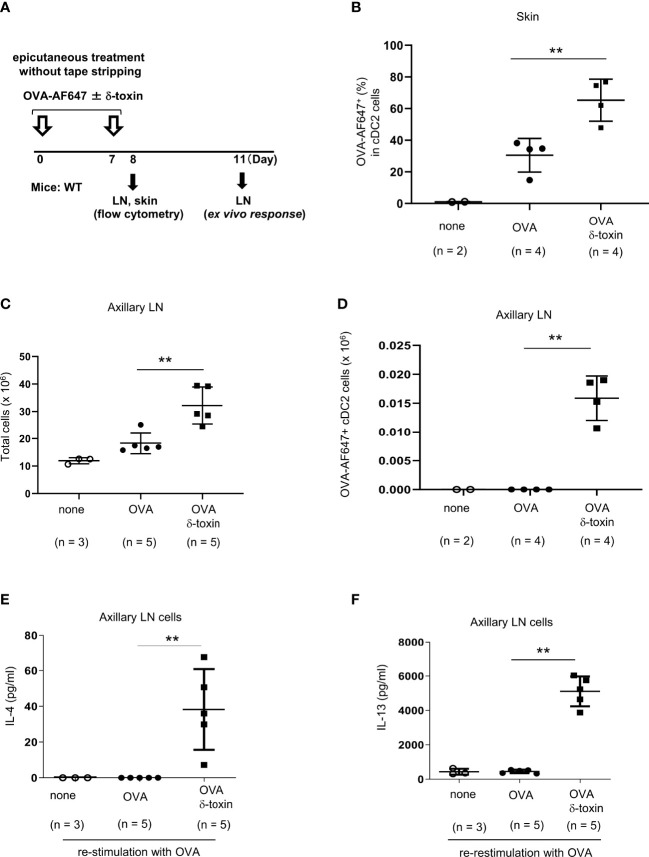
Material and methods: The non-tape-stripped skins of wild-type, KitW-sh/W-sh, or ST2-deficient mice were treated with ovalbumin (OVA) with or without δ-toxin before intragastric administration of OVA. The frequency of diarrhea, numbers of jejunum or skin mast cells, and serum levels of OVA-specific IgE were measured. Conventional dendritic cell 2 (cDC2) in skin and lymph nodes (LN) were analyzed. The cytokine levels in the skin tissues or culture supernatants of δ-toxin-stimulated murine keratinocytes were measured. Anti-IL-1α antibody-pretreated mice were analyzed.
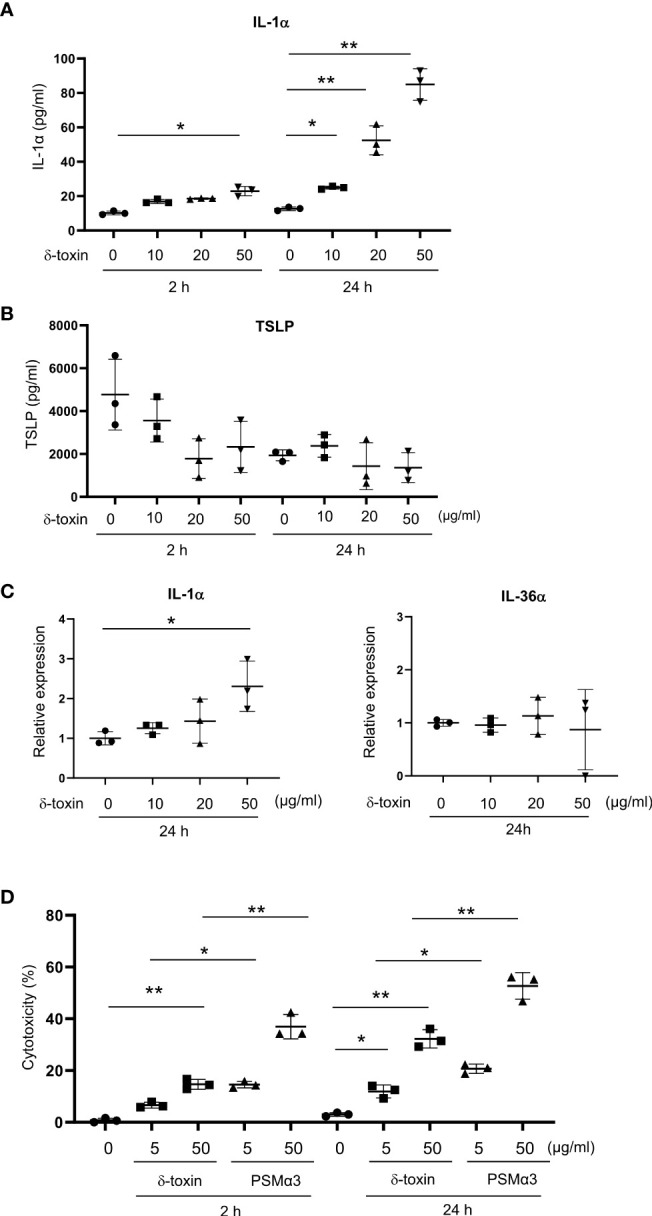
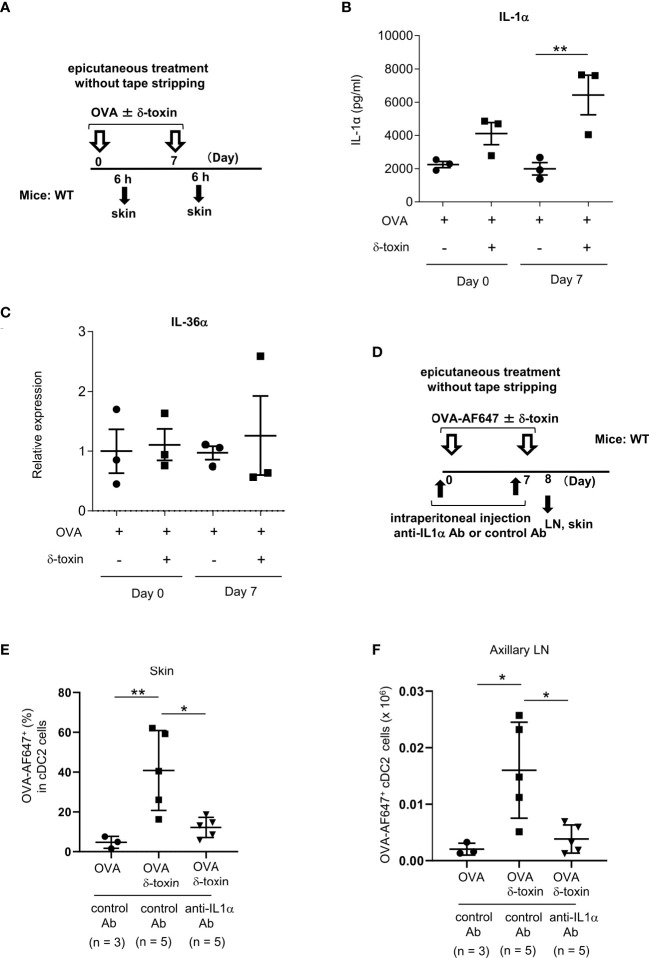
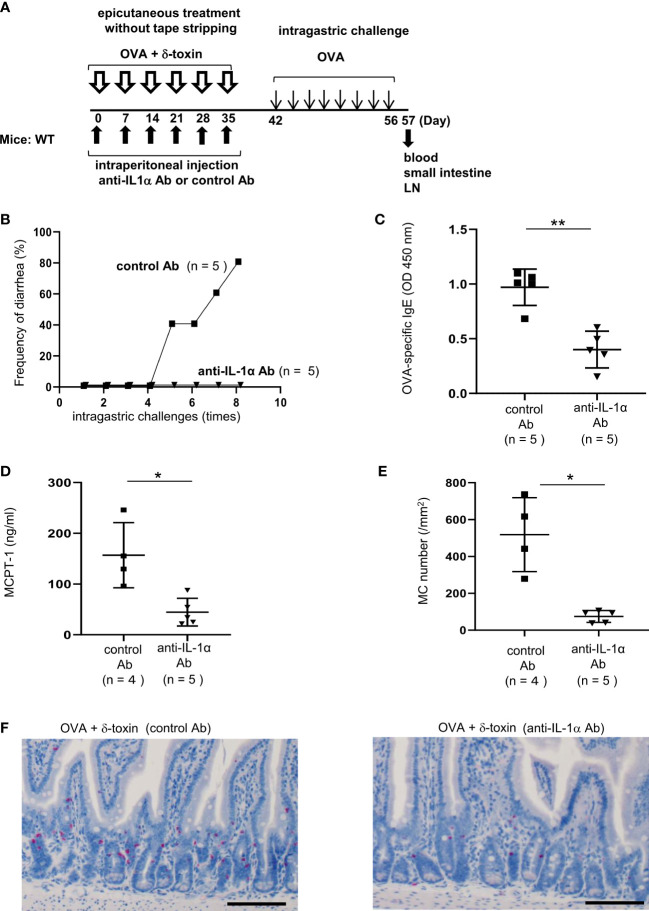
Results: Stimulation with δ-toxin induced the release of IL-1α, but not IL-33, in murine keratinocytes. Epicutaneous treatment with OVA and δ-toxin induced the local production of IL-1α. This treatment induced the translocation of OVA-loaded cDC2 from skin to draining LN and OVA-specific IgE production, independently of mast cells and ST2. This resulted in OVA-administered food allergic responses. In these models, pretreatment with anti-IL-1α antibody inhibited the cDC2 activation and OVA-specific IgE production, thereby dampening food allergic responses.
Conclusion: Even without tape stripping, δ-toxin present on skin enhances epicutaneous sensitization to food allergen in an IL-1α-dependent manner, thereby promoting the development of food allergy.
Keywords: IL-1α; IgE; Staphylococcus aureus δ-toxin; epicutaneous sensitization; food allergy; murine model.
Copyright © 2023 Yamada, Kaitani, Izawa, Ando, Kamei, Uchida, Maehara, Kojima, Yamamoto, Wang, Nagamine, Maeda, Uchida, Nakano, Ohtsuka, Ogawa, Okumura, Shimizu and Kitaura.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous