

A self-transmissible plasmid from a hyperthermophile that facilitates genetic modification of diverse Archaea
- PMID: 37277532
- PMCID: PMC10788138
- DOI: 10.1038/s41564-023-01387-x
A self-transmissible plasmid from a hyperthermophile that facilitates genetic modification of diverse Archaea
Abstract
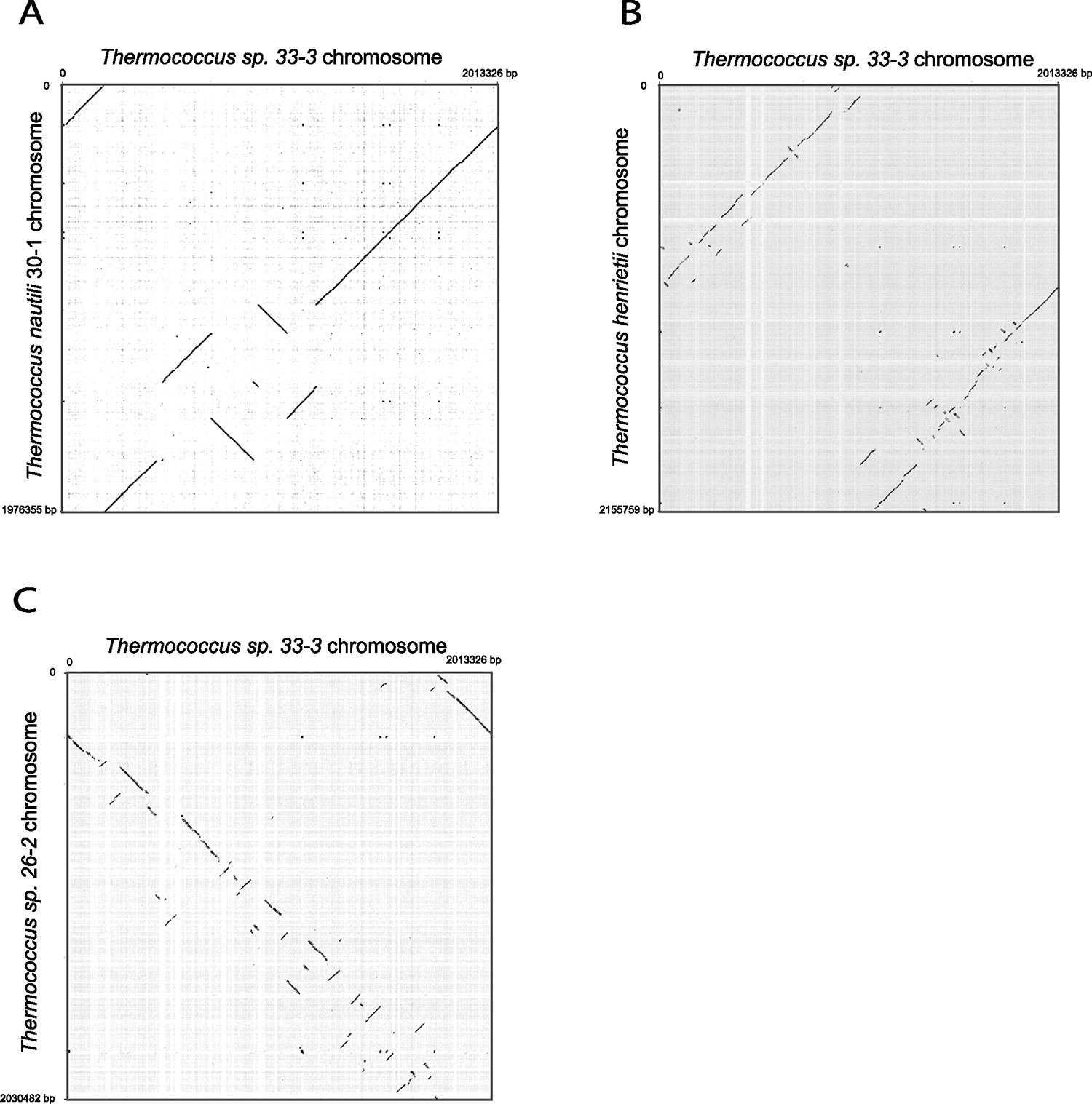
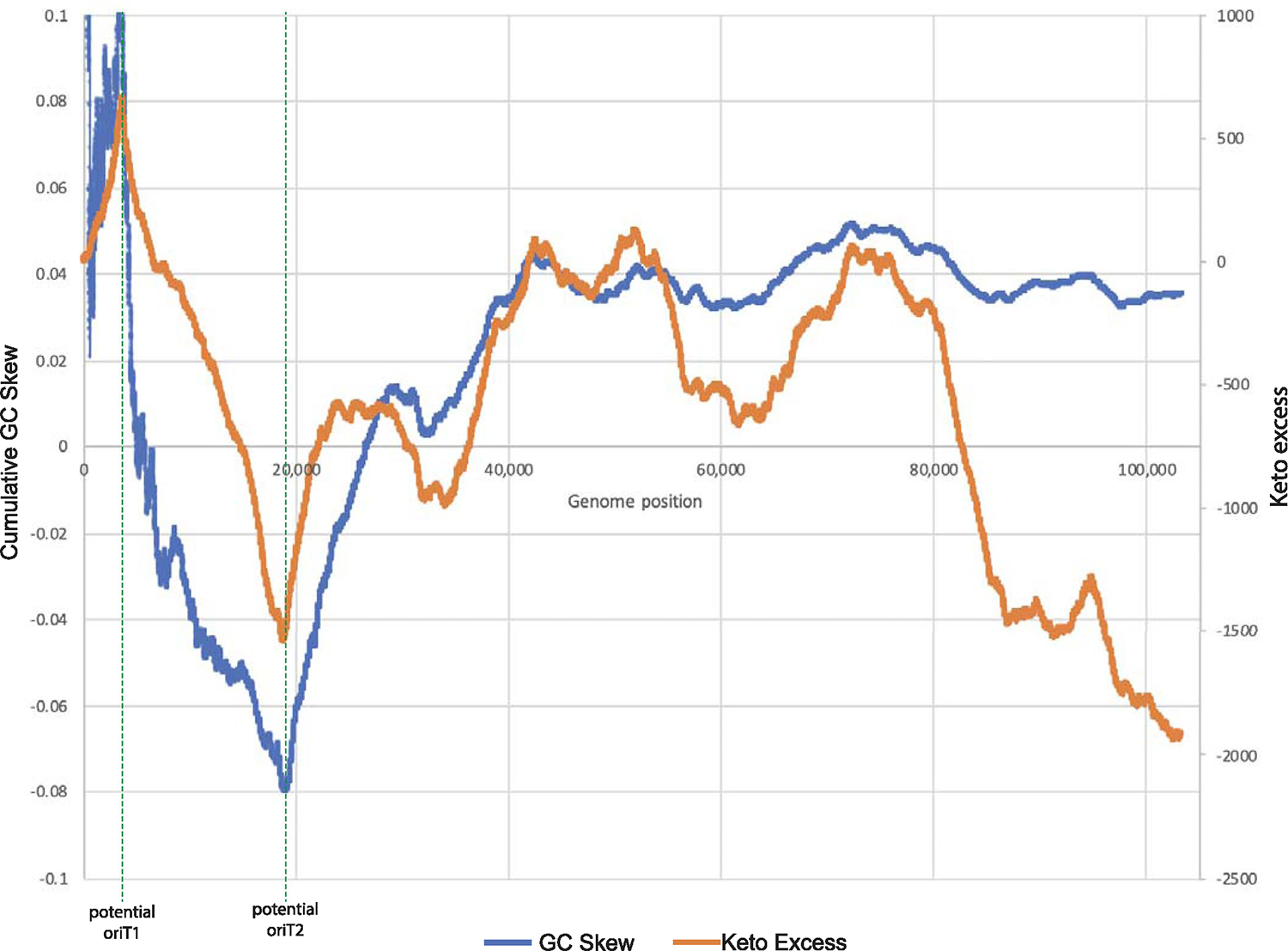
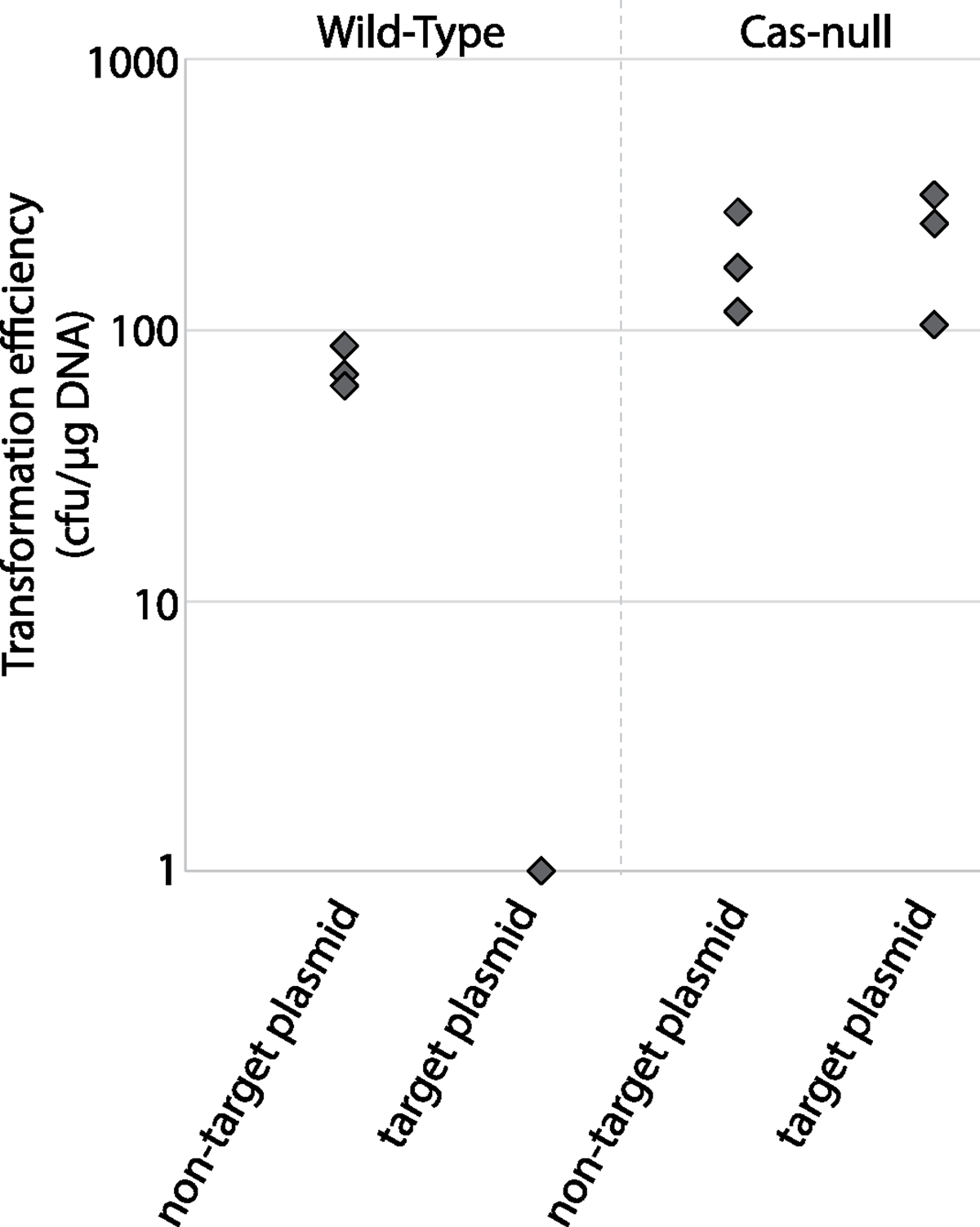
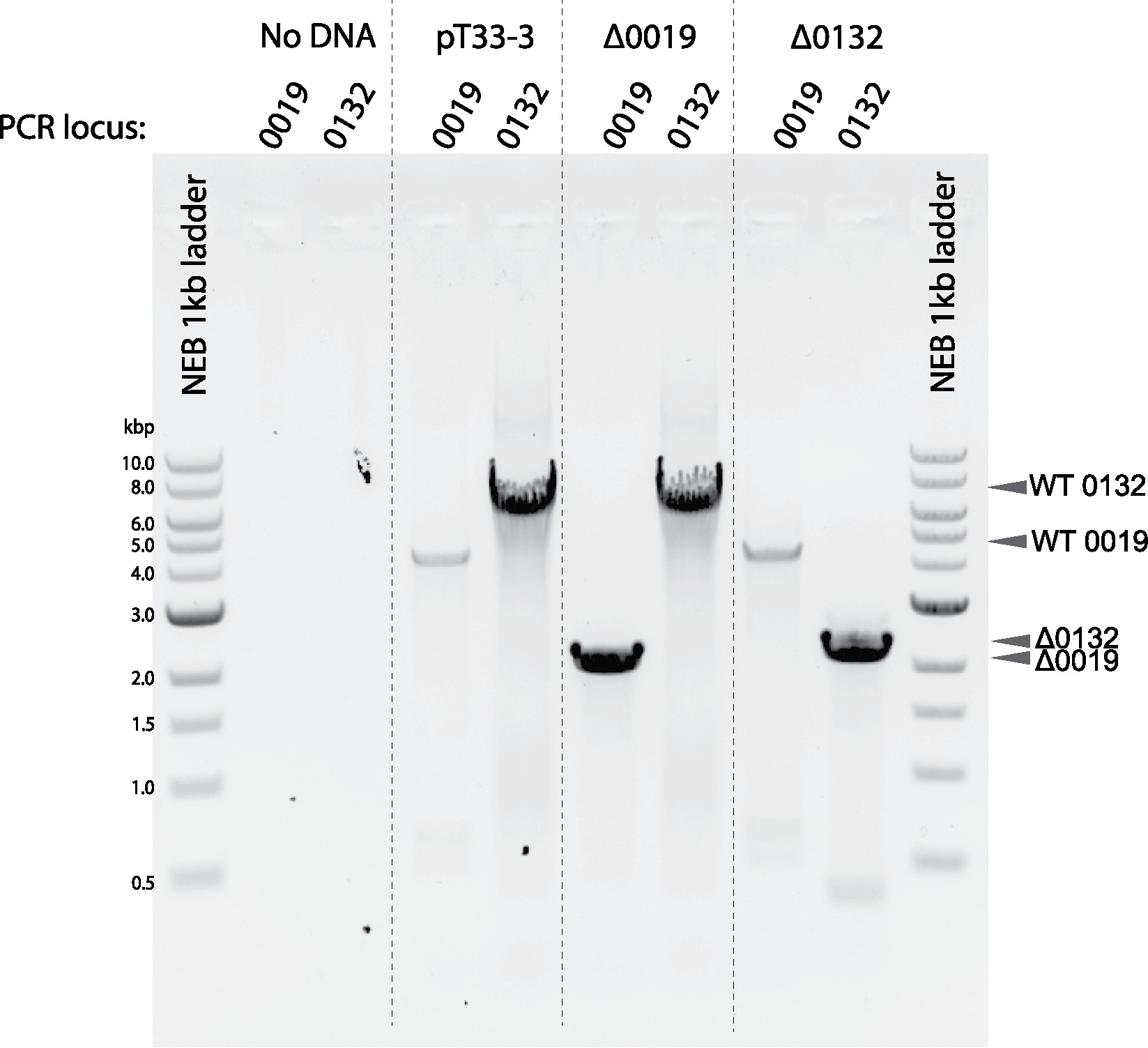
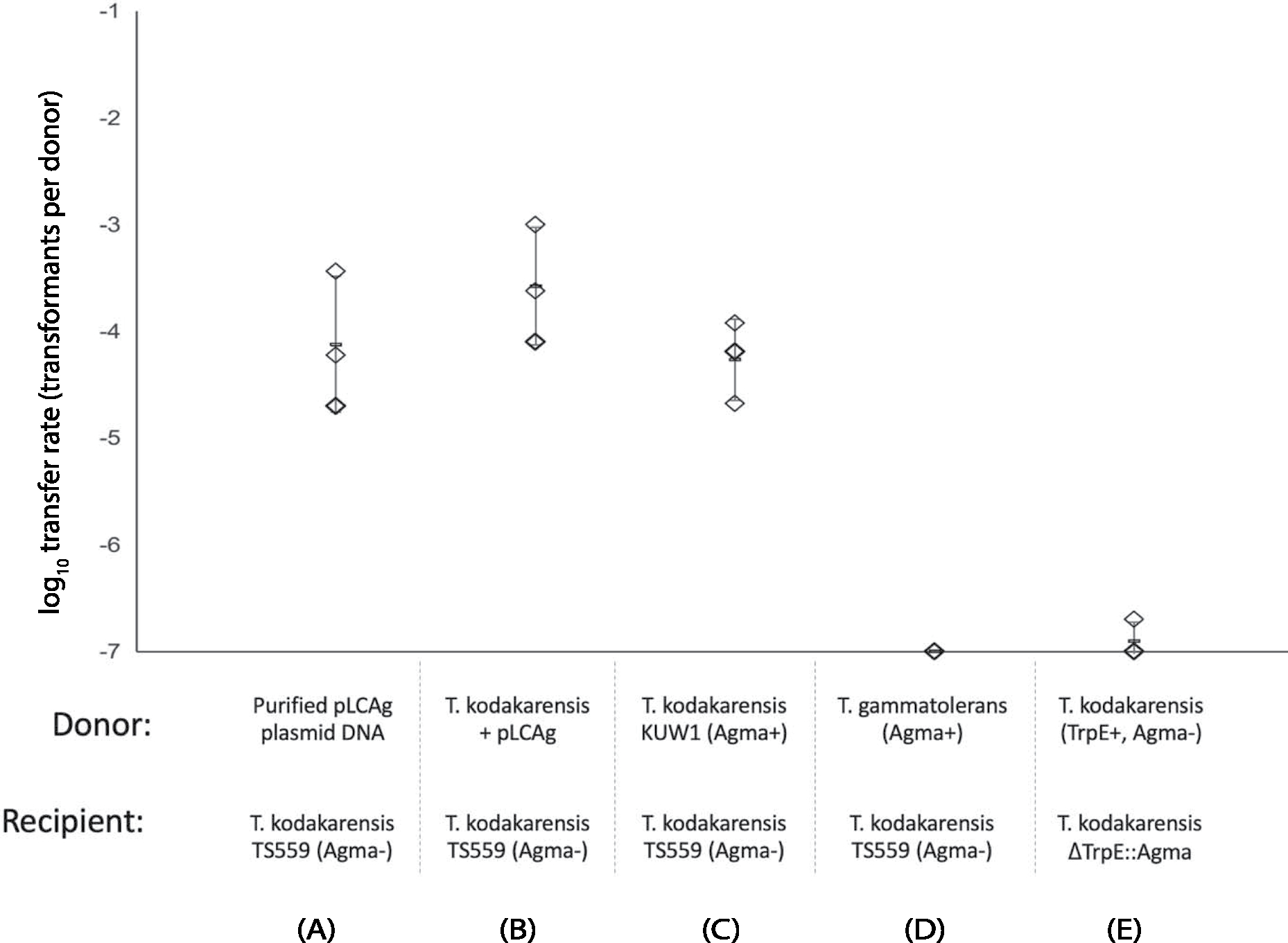
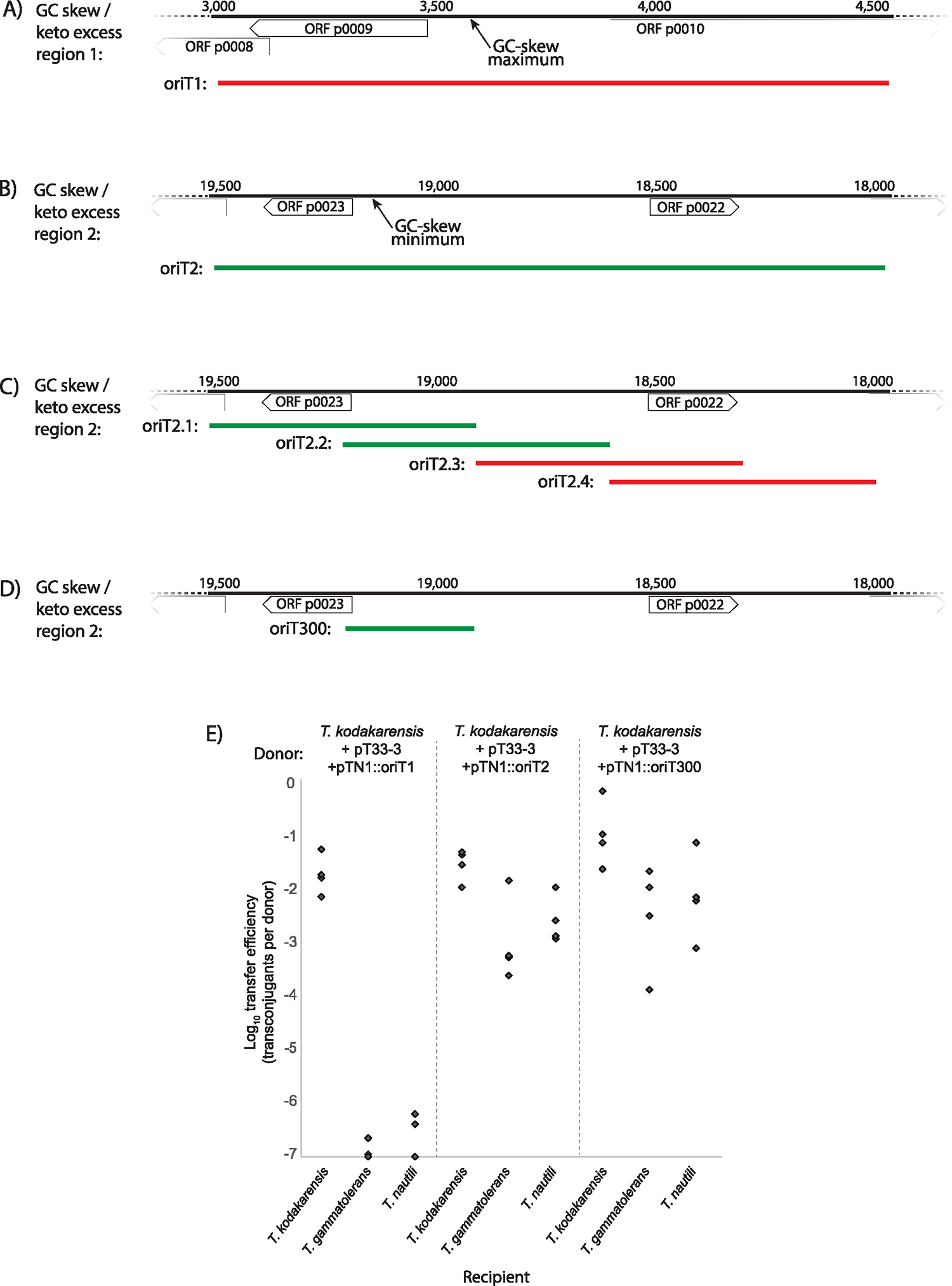
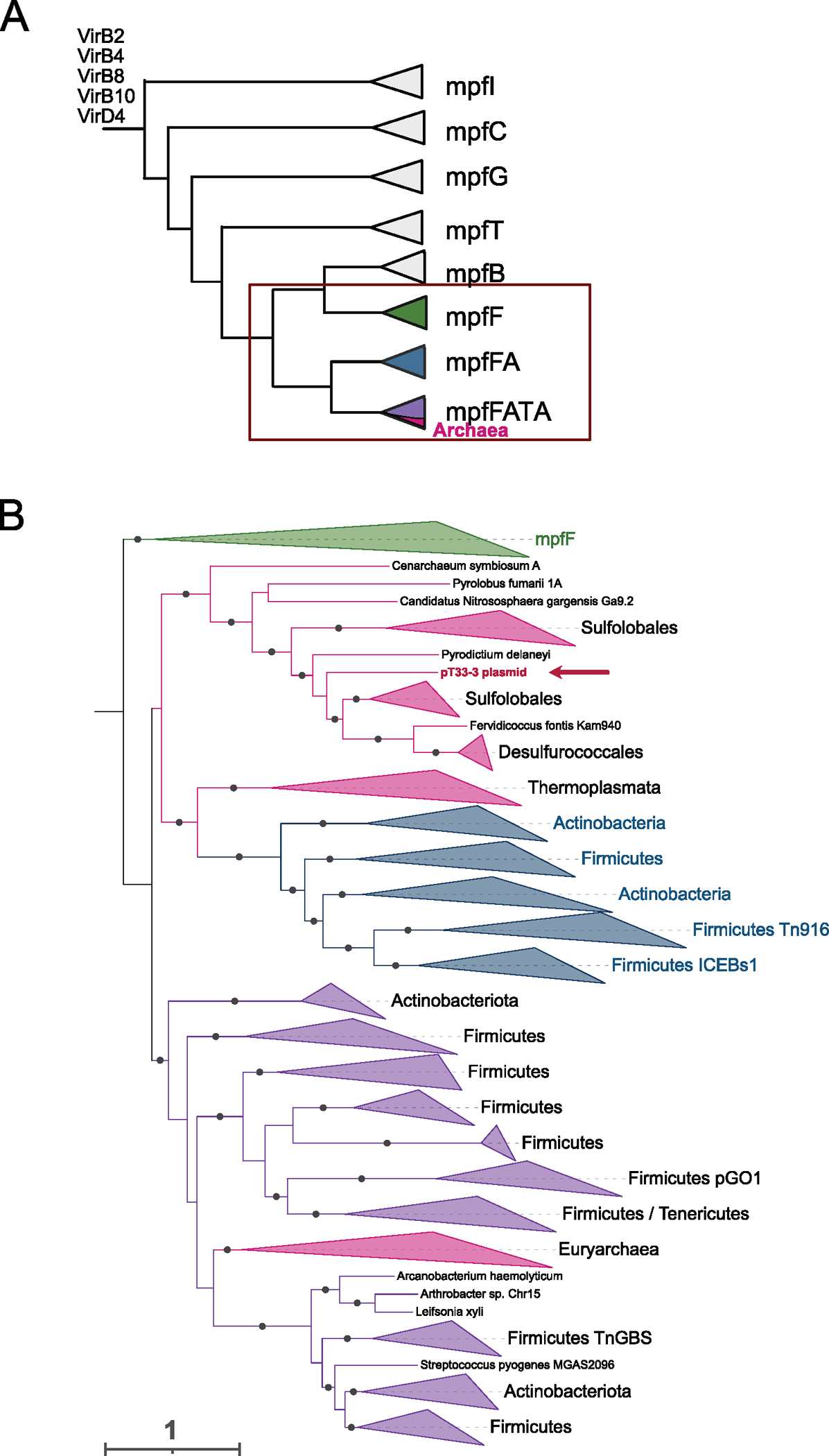
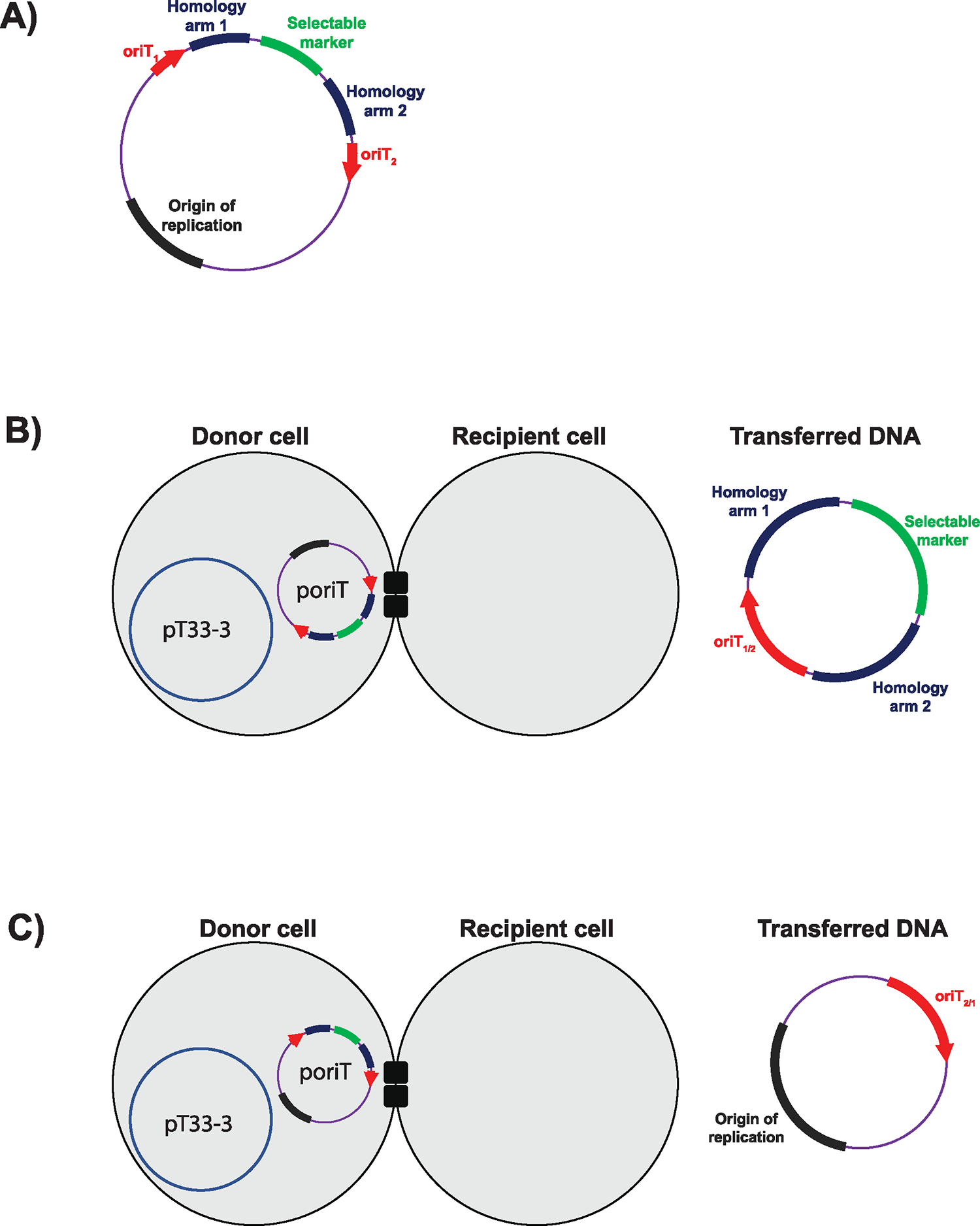
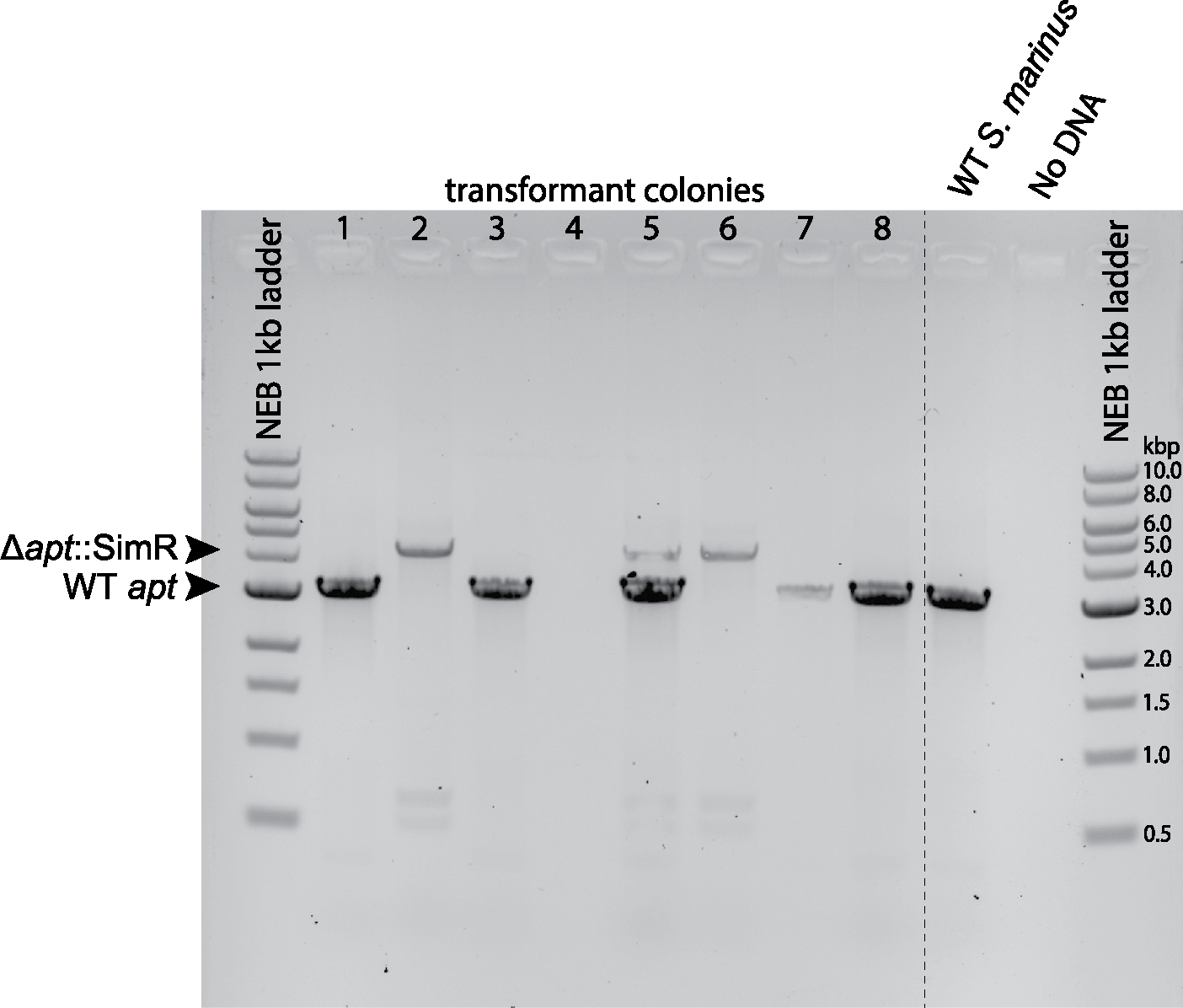
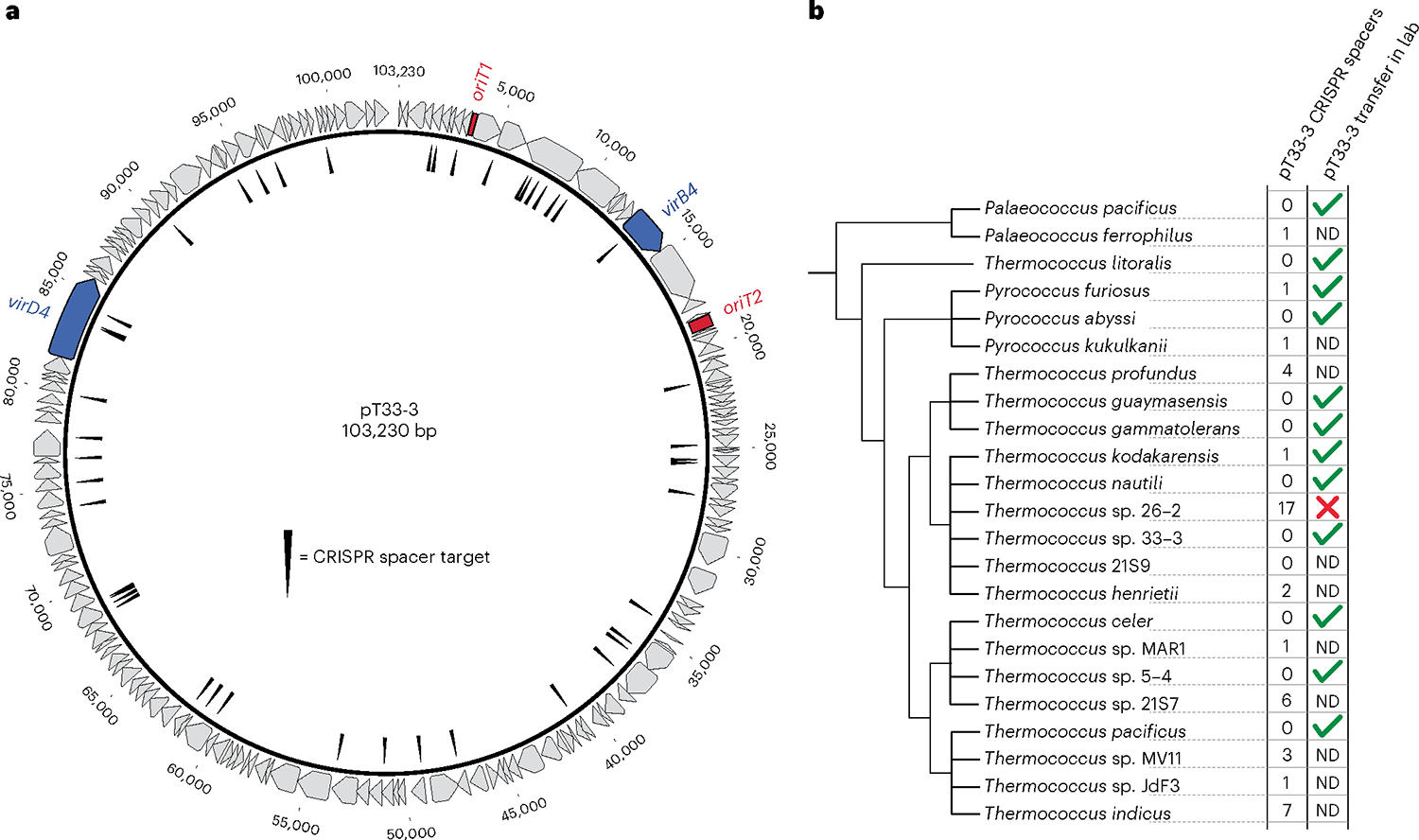
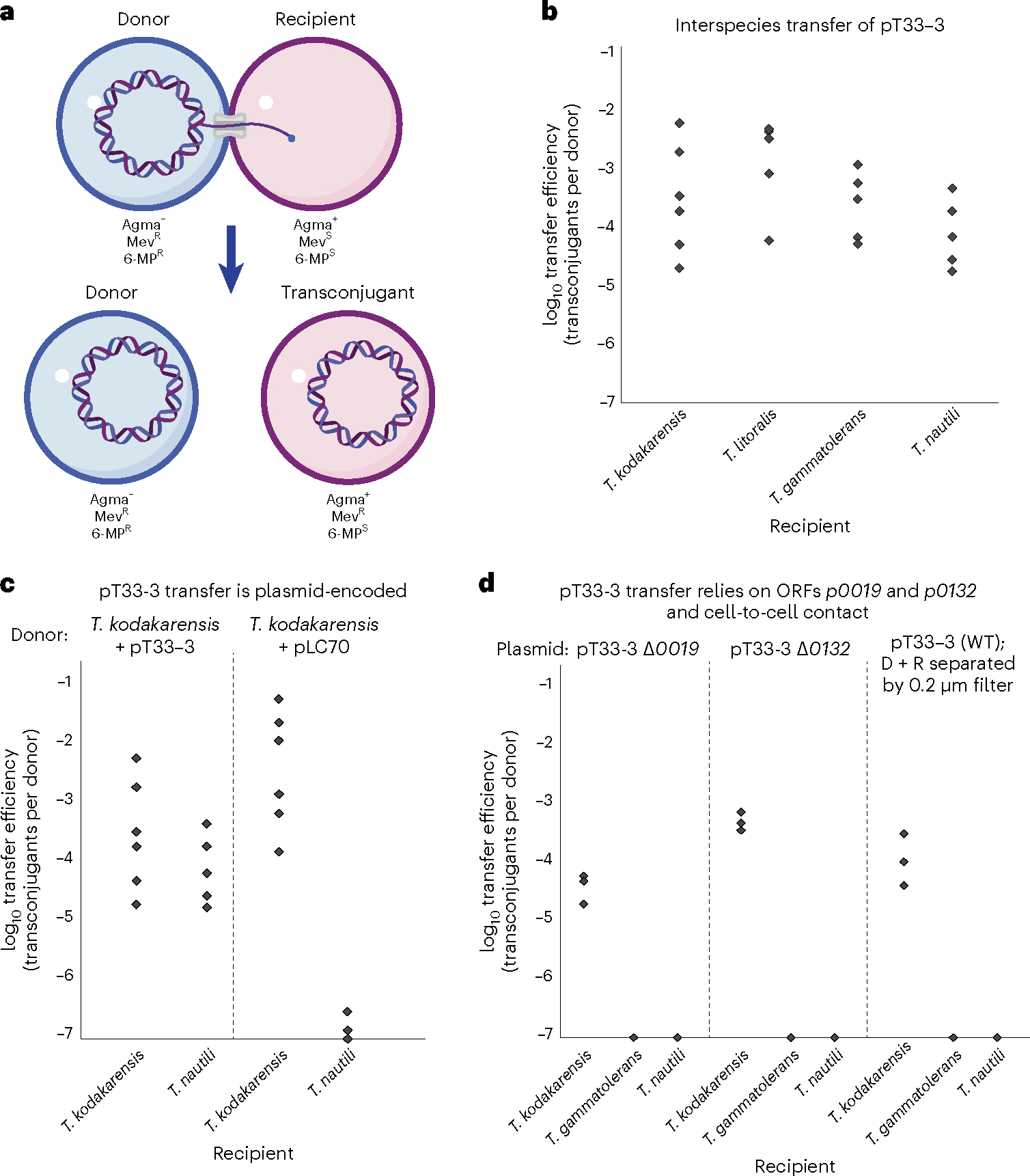
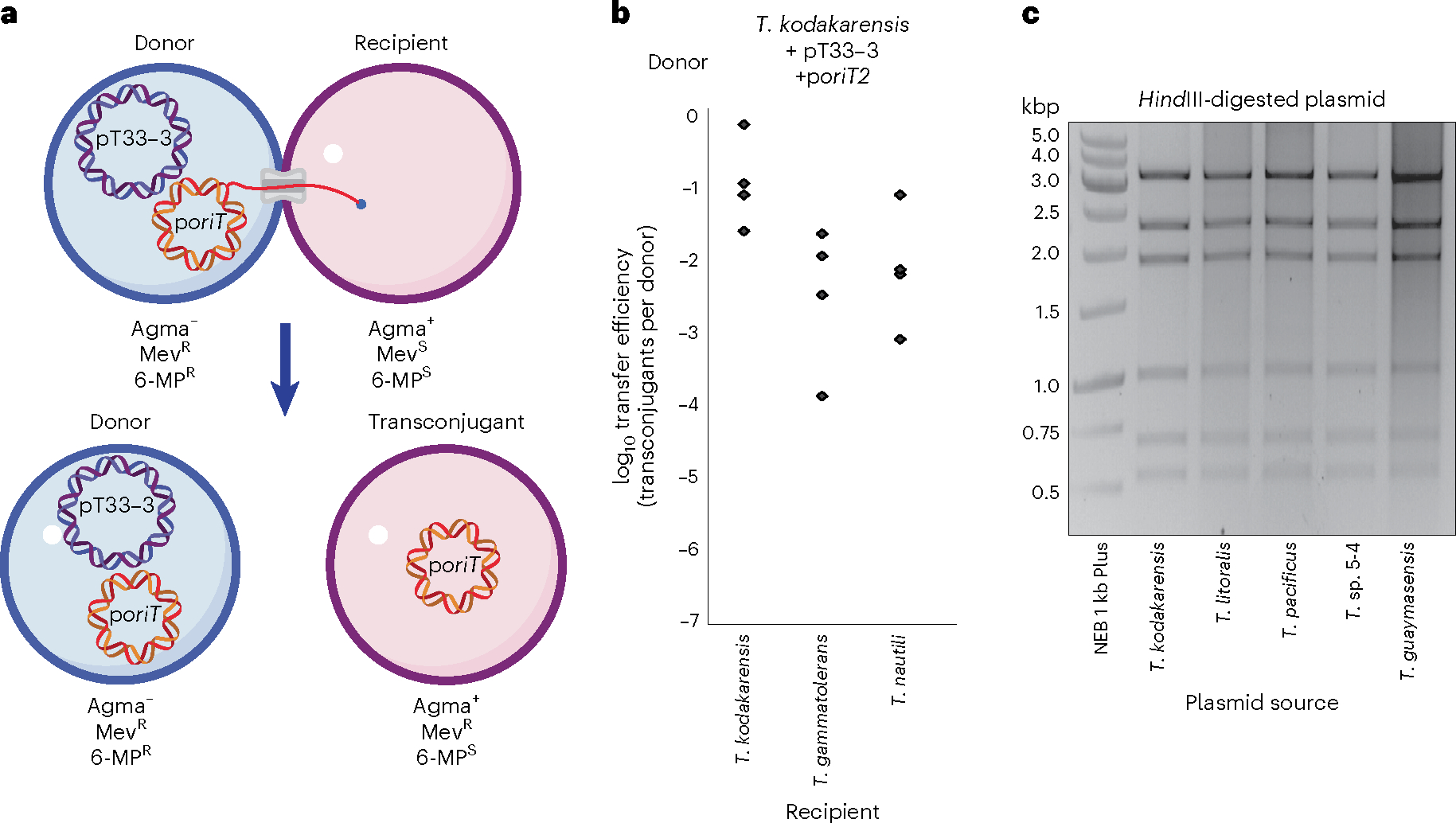
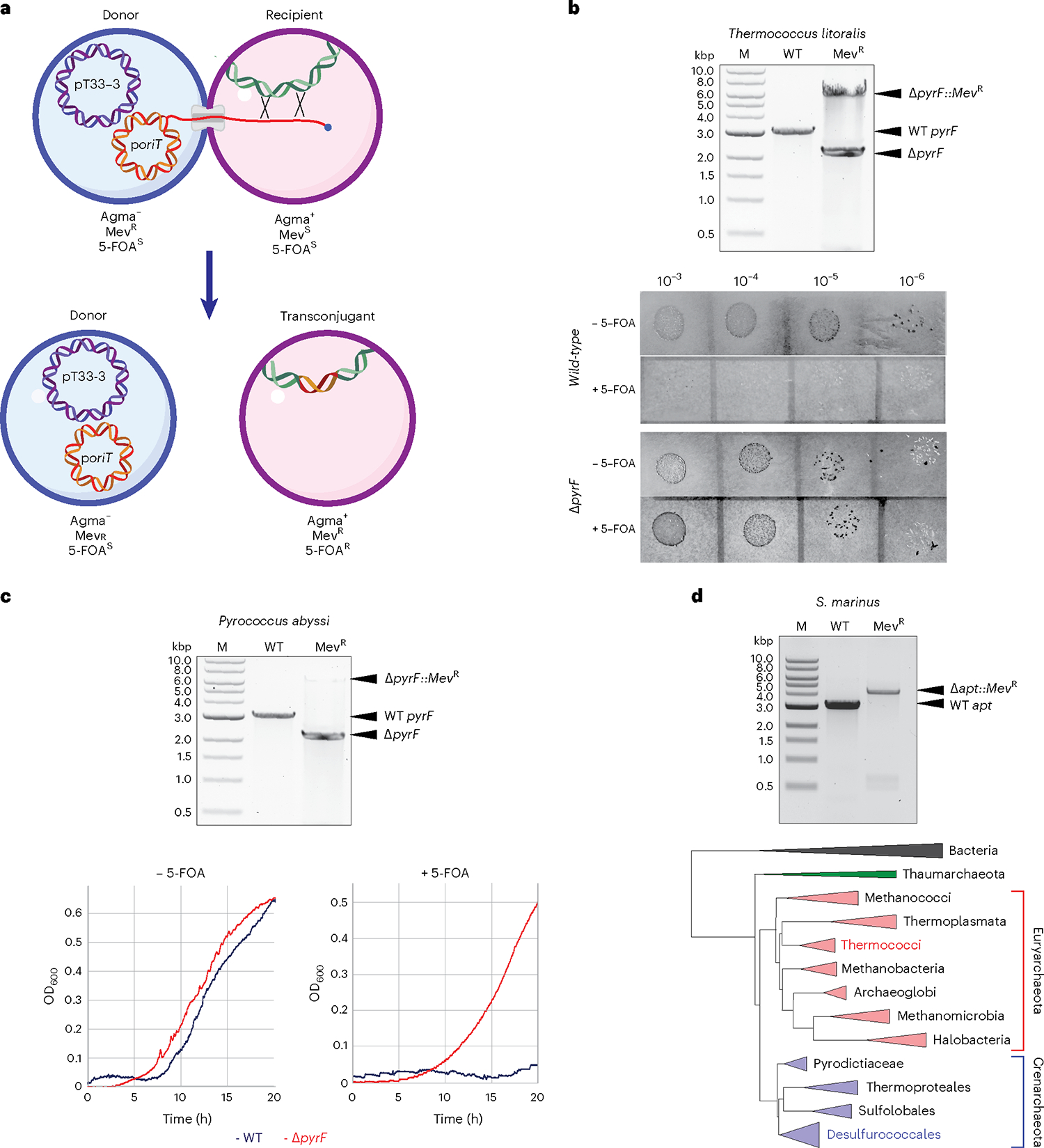
Conjugative plasmids are self-transmissible mobile genetic elements that transfer DNA between host cells via type IV secretion systems (T4SS). While T4SS-mediated conjugation has been well-studied in bacteria, information is sparse in Archaea and known representatives exist only in the Sulfolobales order of Crenarchaeota. Here we present the first self-transmissible plasmid identified in a Euryarchaeon, Thermococcus sp. 33-3. The 103 kbp plasmid, pT33-3, is seen in CRISPR spacers throughout the Thermococcales order. We demonstrate that pT33-3 is a bona fide conjugative plasmid that requires cell-to-cell contact and is dependent on canonical, plasmid-encoded T4SS-like genes. Under laboratory conditions, pT33-3 transfers to various Thermococcales and transconjugants propagate at 100 °C. Using pT33-3, we developed a genetic toolkit that allows modification of phylogenetically diverse Archaeal genomes. We demonstrate pT33-3-mediated plasmid mobilization and subsequent targeted genome modification in previously untransformable Thermococcales species, and extend this process to interphylum transfer to a Crenarchaeon.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
Similar articles
-
Hyper-stimulation of Pyrococcus furiosus CRISPR DNA uptake by a self-transmissible plasmid.Extremophiles. 2022 Nov 16;26(3):36. doi: 10.1007/s00792-022-01281-0. Extremophiles. 2022. PMID: 36385310 Free PMC article.
-
The CRISPR Spacer Space Is Dominated by Sequences from Species-Specific Mobilomes.mBio. 2017 Sep 19;8(5):e01397-17. doi: 10.1128/mBio.01397-17. mBio. 2017. PMID: 28928211 Free PMC article.
-
Protein Transfer through an F Plasmid-Encoded Type IV Secretion System Suppresses the Mating-Induced SOS Response.mBio. 2021 Aug 31;12(4):e0162921. doi: 10.1128/mBio.01629-21. Epub 2021 Jul 13. mBio. 2021. PMID: 34253063 Free PMC article.
-
Driving Apart and Segregating Genomes in Archaea.Trends Microbiol. 2016 Dec;24(12):957-967. doi: 10.1016/j.tim.2016.07.001. Epub 2016 Jul 20. Trends Microbiol. 2016. PMID: 27450111 Free PMC article. Review.
-
Plasmids, viruses and virus-like membrane vesicles from Thermococcales.Biochem Soc Trans. 2011 Jan;39(1):36-44. doi: 10.1042/BST0390036. Biochem Soc Trans. 2011. PMID: 21265744 Review.
Cited by
-
Identification of novel components of the Ced and Ups systems in Saccharolobus islandicus REY15A.mLife. 2025 Feb 23;4(1):17-28. doi: 10.1002/mlf2.12163. eCollection 2025 Feb. mLife. 2025. PMID: 40026581 Free PMC article.
-
A plasmid to modify Archaea.Nat Rev Microbiol. 2023 Aug;21(8):483. doi: 10.1038/s41579-023-00927-w. Nat Rev Microbiol. 2023. PMID: 37322123 No abstract available.
-
The Last Universal Common Ancestor of Ribosome-Encoding Organisms: Portrait of LUCA.J Mol Evol. 2024 Oct;92(5):550-583. doi: 10.1007/s00239-024-10186-9. Epub 2024 Aug 19. J Mol Evol. 2024. PMID: 39158619 Review.
-
Hyper-stimulation of Pyrococcus furiosus CRISPR DNA uptake by a self-transmissible plasmid.Extremophiles. 2022 Nov 16;26(3):36. doi: 10.1007/s00792-022-01281-0. Extremophiles. 2022. PMID: 36385310 Free PMC article.
-
The extended mobility of plasmids.Nucleic Acids Res. 2025 Jul 19;53(14):gkaf652. doi: 10.1093/nar/gkaf652. Nucleic Acids Res. 2025. PMID: 40694848 Free PMC article. Review.
References
-
- de la Cruz F, Frost LS, Meyer RJ & Zechner EL Conjugative DNA metabolism in Gram-negative bacteria. FEMS Microbiol. Rev. 34, 18–40 (2010). - PubMed
-
- Montero I, Herrero-Fresno A, Rodicio R & Rodicio MR Efficient mobilization of a resistance derivative of pSLT, the virulence plasmid specific of Salmonella enterica serovar Typhimurium, by an IncI1 plasmid. Plasmid 70, 104–109 (2013). - PubMed
