

Sexual differentiation in human malaria parasites is regulated by competition between phospholipid metabolism and histone methylation
- PMID: 37277533
- PMCID: PMC11163918
- DOI: 10.1038/s41564-023-01396-w
Sexual differentiation in human malaria parasites is regulated by competition between phospholipid metabolism and histone methylation
Abstract
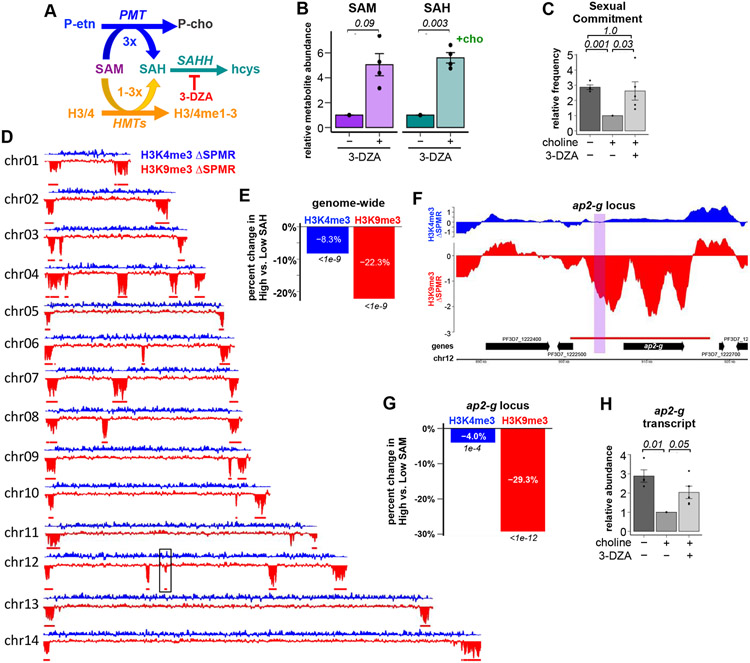
For Plasmodium falciparum, the most widespread and virulent malaria parasite that infects humans, persistence depends on continuous asexual replication in red blood cells, while transmission to their mosquito vector requires asexual blood-stage parasites to differentiate into non-replicating gametocytes. This decision is controlled by stochastic derepression of a heterochromatin-silenced locus encoding AP2-G, the master transcription factor of sexual differentiation. The frequency of ap2-g derepression was shown to be responsive to extracellular phospholipid precursors but the mechanism linking these metabolites to epigenetic regulation of ap2-g was unknown. Through a combination of molecular genetics, metabolomics and chromatin profiling, we show that this response is mediated by metabolic competition for the methyl donor S-adenosylmethionine between histone methyltransferases and phosphoethanolamine methyltransferase, a critical enzyme in the parasite's pathway for de novo phosphatidylcholine synthesis. When phosphatidylcholine precursors are scarce, increased consumption of SAM for de novo phosphatidylcholine synthesis impairs maintenance of the histone methylation responsible for silencing ap2-g, increasing the frequency of derepression and sexual differentiation. This provides a key mechanistic link that explains how LysoPC and choline availability can alter the chromatin status of the ap2-g locus controlling sexual differentiation.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Figures
















References
-
- Drakeley C, Sutherland C, Bousema JT, Sauerwein RW & Targett GAT The epidemiology of Plasmodium falciparum gametocytes: weapons of mass dispersion. Trends in Parasitology 22, 424–430 (2006). - PubMed
-
- Sinha A, Hughes KR, Modrzynska KK, Otto TD, Pfander C, Dickens NJ, Religa AA, Bushell E, Graham AL, Cameron R, Kafsack BFC, Williams AE, Llinás M, Berriman M, Billker O & Waters AP A cascade of DNA-binding proteins for sexual commitment and development in Plasmodium. Nature 507, 253–257 (2014). - PMC - PubMed
-
- Brancucci NMB, Bertschi NL, Zhu L, Niederwieser I, Chin WH, Wampfler R, Freymond C, Rottmann M, Felger I, Bozdech Z & Voss TS Heterochromatin protein 1 secures survival and transmission of malaria parasites. Cell Host Microbe 16, 165–176 (2014). - PubMed
-
- Fraschka SA, Filarsky M, Hoo R, Niederwieser I, Yam XY, Brancucci NMB, Mohring F, Mushunje AT, Huang X, Christensen PR, Nosten F, Bozdech Z, Russell B, Moon RW, Marti M, Preiser PR, Bartfai R & Voss TS Comparative Heterochromatin Profiling Reveals Conserved and Unique Epigenome Signatures Linked to Adaptation and Development of Malaria Parasites. Cell Host Microbe 23, 407–420.e8 (2018). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
