

Chromatin alternates between A and B compartments at kilobase scale for subgenic organization
- PMID: 37280210
- PMCID: PMC10244318
- DOI: 10.1038/s41467-023-38429-1
Chromatin alternates between A and B compartments at kilobase scale for subgenic organization
Abstract
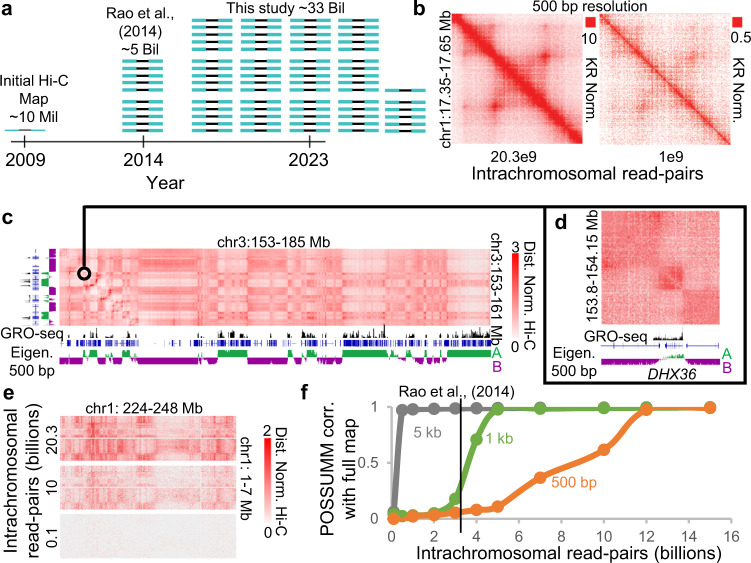
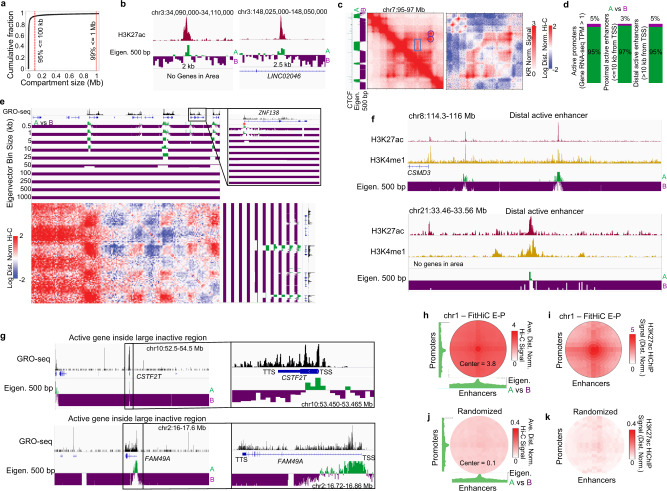
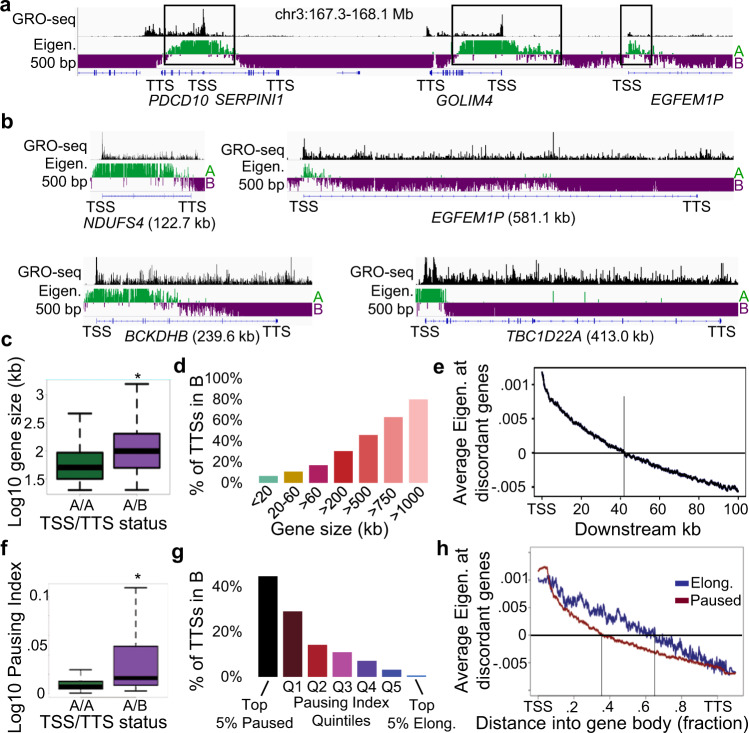
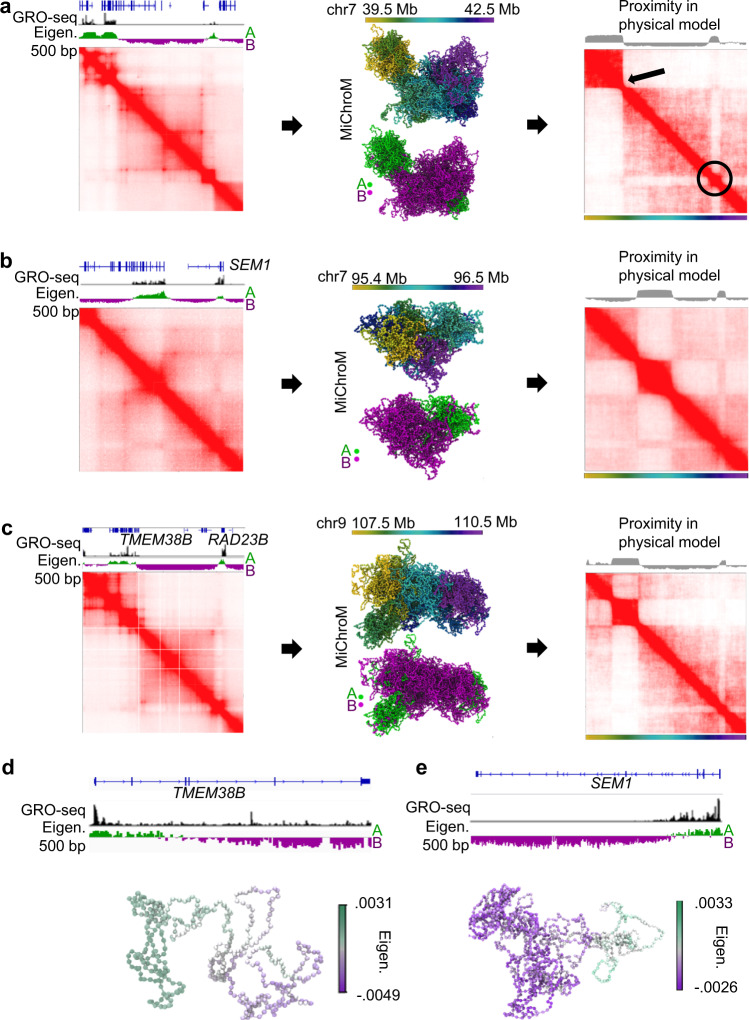
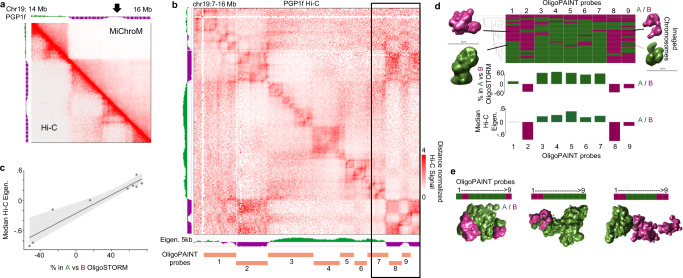
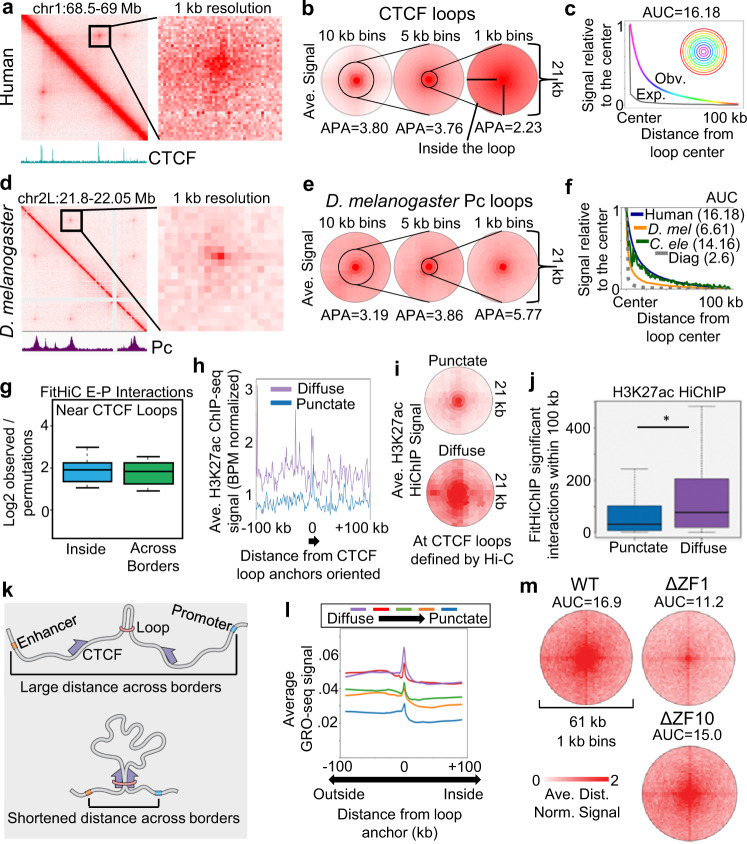
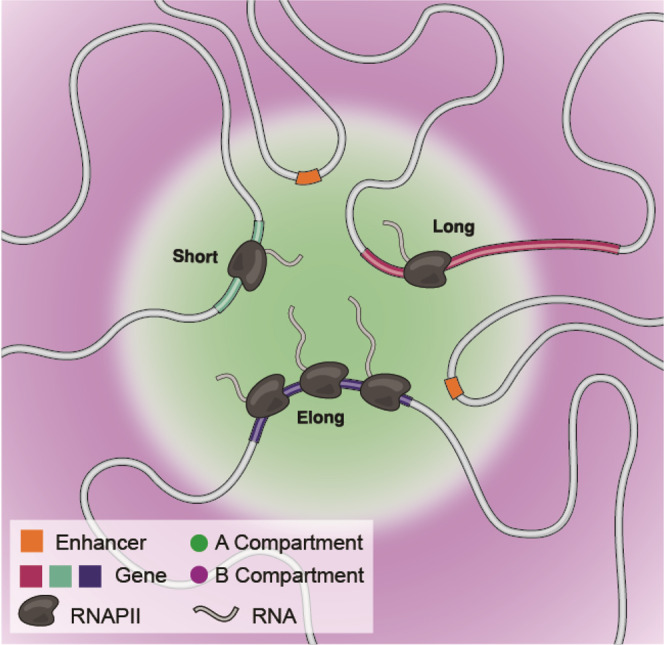
Nuclear compartments are prominent features of 3D chromatin organization, but sequencing depth limitations have impeded investigation at ultra fine-scale. CTCF loops are generally studied at a finer scale, but the impact of looping on proximal interactions remains enigmatic. Here, we critically examine nuclear compartments and CTCF loop-proximal interactions using a combination of in situ Hi-C at unparalleled depth, algorithm development, and biophysical modeling. Producing a large Hi-C map with 33 billion contacts in conjunction with an algorithm for performing principal component analysis on sparse, super massive matrices (POSSUMM), we resolve compartments to 500 bp. Our results demonstrate that essentially all active promoters and distal enhancers localize in the A compartment, even when flanking sequences do not. Furthermore, we find that the TSS and TTS of paused genes are often segregated into separate compartments. We then identify diffuse interactions that radiate from CTCF loop anchors, which correlate with strong enhancer-promoter interactions and proximal transcription. We also find that these diffuse interactions depend on CTCF's RNA binding domains. In this work, we demonstrate features of fine-scale chromatin organization consistent with a revised model in which compartments are more precise than commonly thought while CTCF loops are more protracted.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Acute depletion of CTCF directly affects MYC regulation through loss of enhancer-promoter looping.Nucleic Acids Res. 2019 Jul 26;47(13):6699-6713. doi: 10.1093/nar/gkz462. Nucleic Acids Res. 2019. PMID: 31127282 Free PMC article.
-
Interplay between CTCF boundaries and a super enhancer controls cohesin extrusion trajectories and gene expression.Mol Cell. 2021 Aug 5;81(15):3082-3095.e6. doi: 10.1016/j.molcel.2021.06.008. Epub 2021 Jun 30. Mol Cell. 2021. PMID: 34197738
-
Three-dimensional genome architectural CCCTC-binding factor makes choice in duplicated enhancers at Pcdhα locus.Sci China Life Sci. 2020 Jun;63(6):835-844. doi: 10.1007/s11427-019-1598-4. Epub 2020 Apr 2. Sci China Life Sci. 2020. PMID: 32249388
-
CTCF: the protein, the binding partners, the binding sites and their chromatin loops.Philos Trans R Soc Lond B Biol Sci. 2013 May 6;368(1620):20120369. doi: 10.1098/rstb.2012.0369. Print 2013. Philos Trans R Soc Lond B Biol Sci. 2013. PMID: 23650640 Free PMC article. Review.
-
Dissecting CTCF site function in a tense HoxD locus.Genes Dev. 2021 Nov 1;35(21-22):1401-1402. doi: 10.1101/gad.349089.121. Genes Dev. 2021. PMID: 34725128 Free PMC article. Review.
Cited by
-
Machine and Deep Learning Methods for Predicting 3D Genome Organization.Methods Mol Biol. 2025;2856:357-400. doi: 10.1007/978-1-0716-4136-1_22. Methods Mol Biol. 2025. PMID: 39283464 Review.
-
LAP2alpha facilitates myogenic gene expression by preventing nucleoplasmic lamin A/C from spreading to active chromatin regions.Nucleic Acids Res. 2024 Oct 28;52(19):11500-11518. doi: 10.1093/nar/gkae752. Nucleic Acids Res. 2024. PMID: 39228367 Free PMC article.
-
Genome structure mapping with high-resolution 3D genomics and deep learning.bioRxiv [Preprint]. 2025 May 7:2025.05.06.650874. doi: 10.1101/2025.05.06.650874. bioRxiv. 2025. PMID: 40654659 Free PMC article. Preprint.
-
Leveraging Hi-C Data to Detect Chromosomal Reorganizations.Methods Mol Biol. 2025;2968:213-230. doi: 10.1007/978-1-0716-4750-9_12. Methods Mol Biol. 2025. PMID: 40884646
-
Machine learning tools for deciphering the regulatory logic of enhancers in health and disease.Front Genet. 2025 Aug 13;16:1603687. doi: 10.3389/fgene.2025.1603687. eCollection 2025. Front Genet. 2025. PMID: 40881170 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
