

Molecular basis for the recognition of 24-(S)-hydroxycholesterol by integrin αvβ3
- PMID: 37280310
- PMCID: PMC10244445
- DOI: 10.1038/s41598-023-36040-4
Molecular basis for the recognition of 24-(S)-hydroxycholesterol by integrin αvβ3
Abstract
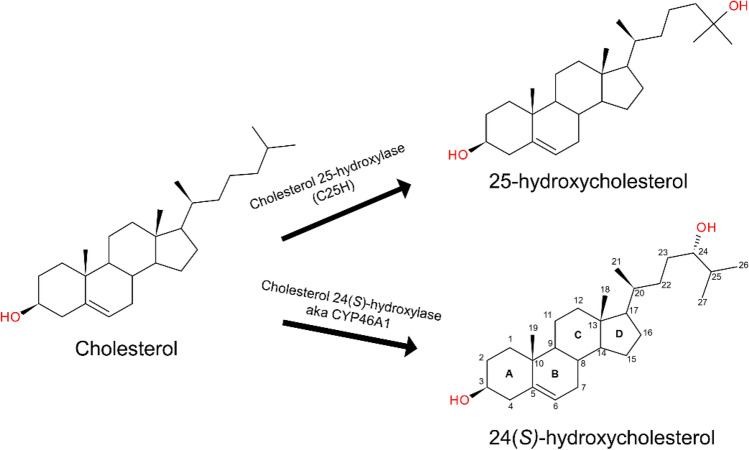
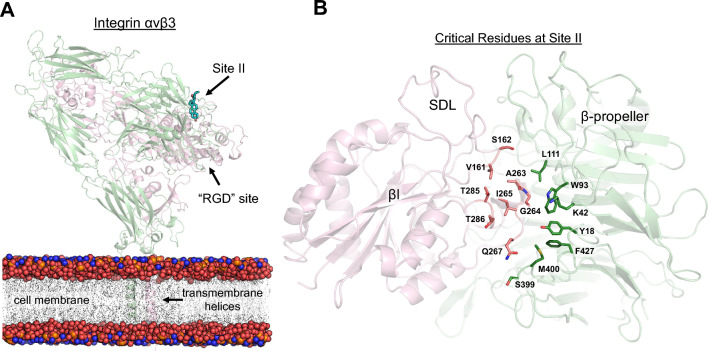
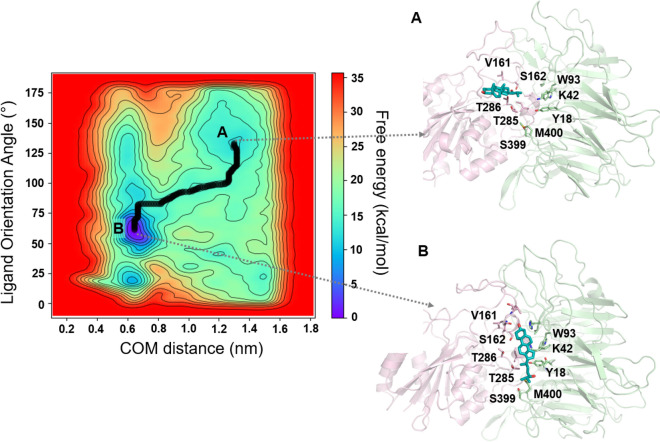
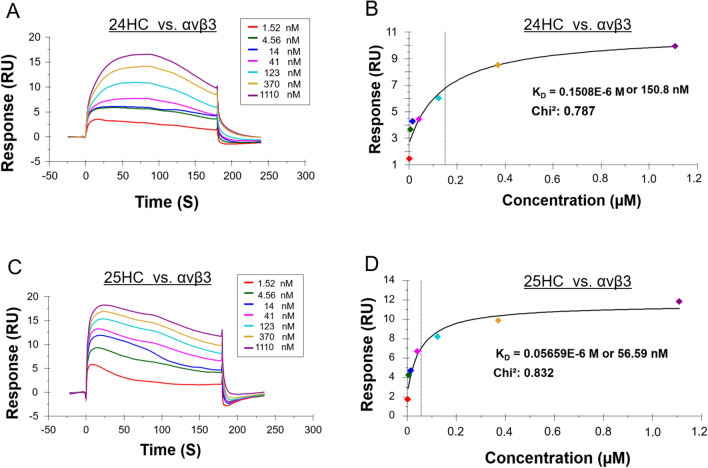
A growing body of evidence suggests that oxysterols such as 25-hydroxycholesterol (25HC) are biologically active and involved in many physiological and pathological processes. Our previous study demonstrated that 25HC induces an innate immune response during viral infections by activating the integrin-focal adhesion kinase (FAK) pathway. 25HC produced the proinflammatory response by binding directly to integrins at a novel binding site (site II) and triggering the production of proinflammatory mediators such as tumor necrosis factor-α (TNF) and interleukin-6 (IL-6). 24-(S)-hydroxycholesterol (24HC), a structural isomer of 25HC, plays a critical role in cholesterol homeostasis in the human brain and is implicated in multiple inflammatory conditions, including Alzheimer's disease. However, whether 24HC can induce a proinflammatory response like 25HC in non-neuronal cells has not been studied and remains unknown. The aim of this study was to examine whether 24HC produces such an immune response using in silico and in vitro experiments. Our results indicate that despite being a structural isomer of 25HC, 24HC binds at site II in a distinct binding mode, engages in varied residue interactions, and produces significant conformational changes in the specificity-determining loop (SDL). In addition, our surface plasmon resonance (SPR) study reveals that 24HC could directly bind to integrin αvβ3, with a binding affinity three-fold lower than 25HC. Furthermore, our in vitro studies with macrophages support the involvement of FAK and NFκB signaling pathways in triggering 24HC-mediated production of TNF. Thus, we have identified 24HC as another oxysterol that binds to integrin αvβ3 and promotes a proinflammatory response via the integrin-FAK-NFκB pathway.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2017 Dec 22;12(12):CD011535. doi: 10.1002/14651858.CD011535.pub2. Cochrane Database Syst Rev. 2017. Update in: Cochrane Database Syst Rev. 2020 Jan 9;1:CD011535. doi: 10.1002/14651858.CD011535.pub3. PMID: 29271481 Free PMC article. Updated.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2021 Apr 19;4(4):CD011535. doi: 10.1002/14651858.CD011535.pub4. Cochrane Database Syst Rev. 2021. Update in: Cochrane Database Syst Rev. 2022 May 23;5:CD011535. doi: 10.1002/14651858.CD011535.pub5. PMID: 33871055 Free PMC article. Updated.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2020 Jan 9;1(1):CD011535. doi: 10.1002/14651858.CD011535.pub3. Cochrane Database Syst Rev. 2020. Update in: Cochrane Database Syst Rev. 2021 Apr 19;4:CD011535. doi: 10.1002/14651858.CD011535.pub4. PMID: 31917873 Free PMC article. Updated.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
Cited by
-
Phase 1 pharmacokinetic and safety study of soticlestat in participants with mild or moderate hepatic impairment or normal hepatic function.Pharmacol Res Perspect. 2024 Aug;12(4):e1213. doi: 10.1002/prp2.1213. Pharmacol Res Perspect. 2024. PMID: 38993008 Free PMC article. Clinical Trial.
-
Immune Implications of Cholesterol-Containing Lipid Nanoparticles.ACS Nano. 2024 Oct 22;18(42):28480-28501. doi: 10.1021/acsnano.4c06369. Epub 2024 Oct 10. ACS Nano. 2024. PMID: 39388645 Review.
-
Bottlenecks in the Investigation of Retinal Sterol Homeostasis.Biomolecules. 2024 Mar 12;14(3):341. doi: 10.3390/biom14030341. Biomolecules. 2024. PMID: 38540760 Free PMC article. Review.
-
Increased 25-hydroxycholesterol as an indicator for patients with vestibular neuritis.Front Neurol. 2025 Jun 20;16:1600185. doi: 10.3389/fneur.2025.1600185. eCollection 2025. Front Neurol. 2025. PMID: 40621094 Free PMC article.
-
The mechanism of 25-hydroxycholesterol-mediated suppression of atrial β1-adrenergic responses.Pflugers Arch. 2024 Mar;476(3):407-421. doi: 10.1007/s00424-024-02913-4. Epub 2024 Jan 22. Pflugers Arch. 2024. PMID: 38253680
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
