

Integrated multi-omics analyses reveal homology-directed repair pathway as a unique dependency in near-haploid leukemia
- PMID: 37286545
- PMCID: PMC10247733
- DOI: 10.1038/s41408-023-00863-1
Integrated multi-omics analyses reveal homology-directed repair pathway as a unique dependency in near-haploid leukemia
Abstract
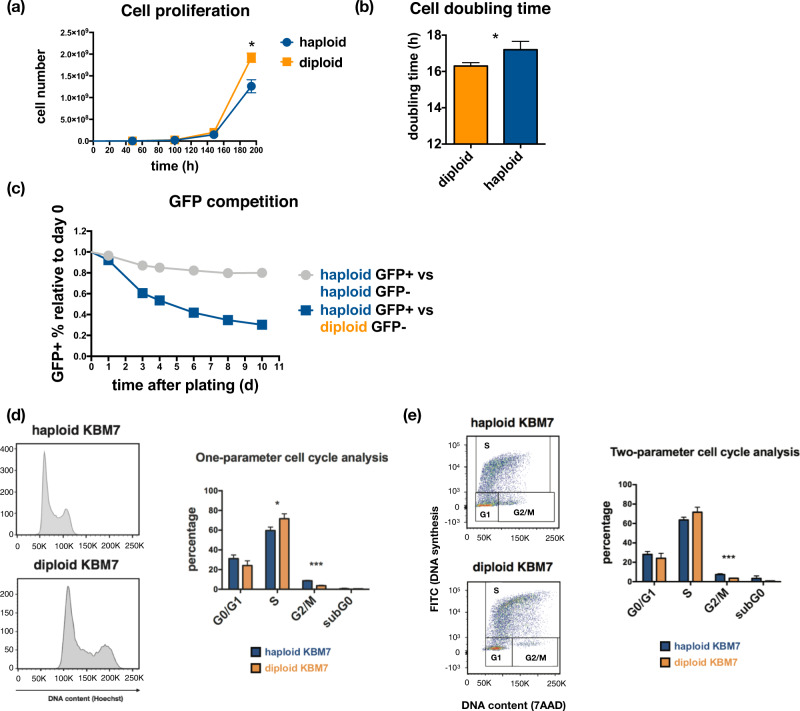
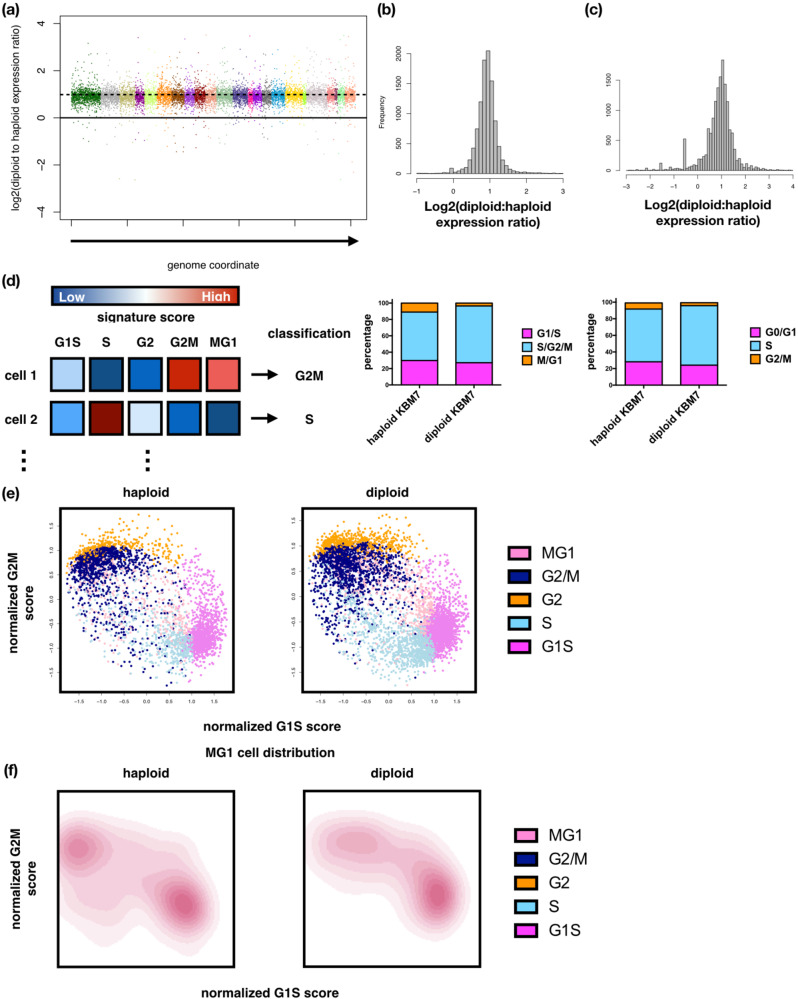
Whole chromosome losses resulting in near-haploid karyotypes are found in a rare subgroup of treatment-refractory acute lymphoblastic leukemia. To systematically dissect the unique physiology and uncover susceptibilities that can be exploited in near-haploid leukemia, we leveraged single-cell RNA-Seq and computational inference of cell cycle stages to pinpoint key differences between near-haploid and diploid leukemia cells. Combining cell cycle stage-specific differential expression with gene essentiality scores from a genome-wide CRISPR-Cas9-mediated knockout screen, we identified the homologous recombination pathway component RAD51B as an essential gene in near-haploid leukemia. DNA damage analyses revealed significantly increased sensitivity of RAD51-mediated repair to RAD51B loss in the G2/M stage of near-haploid cells, suggesting a unique role of RAD51B in the homologous recombination pathway. Elevated G2/M and G1/S checkpoint signaling was part of a RAD51B signature expression program in response to chemotherapy in a xenograft model of human near-haploid B-ALL, and RAD51B and its associated programs were overexpressed in a large panel of near-haploid B-ALL patients. These data highlight a unique genetic dependency on DNA repair machinery in near-haploid leukemia and demarcate RAD51B as a promising candidate for targeted therapy in this treatment-resistant disease.
© 2023. The Author(s).
Conflict of interest statement
D.M.W. is an employee of Merck Research Laboratories. A.R. is a co-founder and equity holder of Celsius Therapeutics, an equity holder in Immunitas, and until July 31, 2020 was an SAB member of ThermoFisher Scientific, Syros Pharmaceuticals, Neogene Therapeutics and Asimov. From August 1, 2020, A.R. is an employee of Genentech, and has equity in Roche.
Figures




References
-
- Ries LAG, Smith MA, Gurney JG, Linet M, Tamra T, Young JL, et al. Cancer incidence and survival among children and adolescents: United States SEER Program 1975–1995. NIH Pub No 99-4649. 1999; 179 pp.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
