

Evaluating the mouse neural precursor line, SN4741, as a suitable proxy for midbrain dopaminergic neurons
- PMID: 37286935
- PMCID: PMC10245633
- DOI: 10.1186/s12864-023-09398-y
Evaluating the mouse neural precursor line, SN4741, as a suitable proxy for midbrain dopaminergic neurons
Abstract
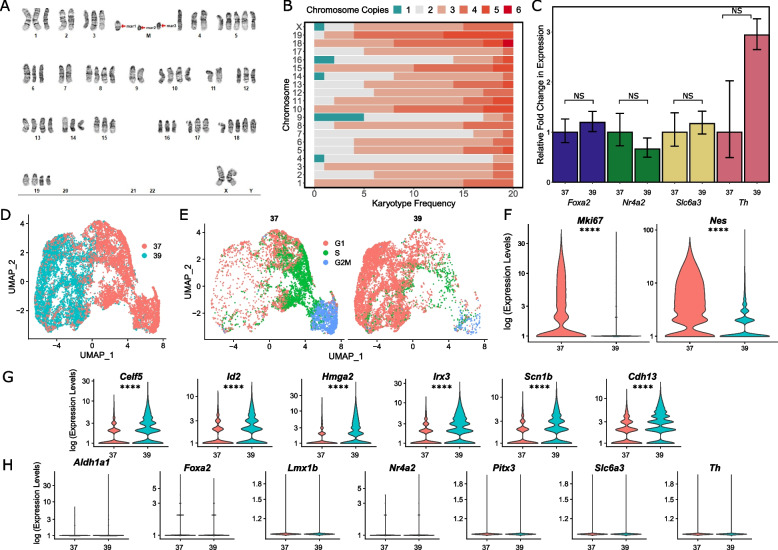
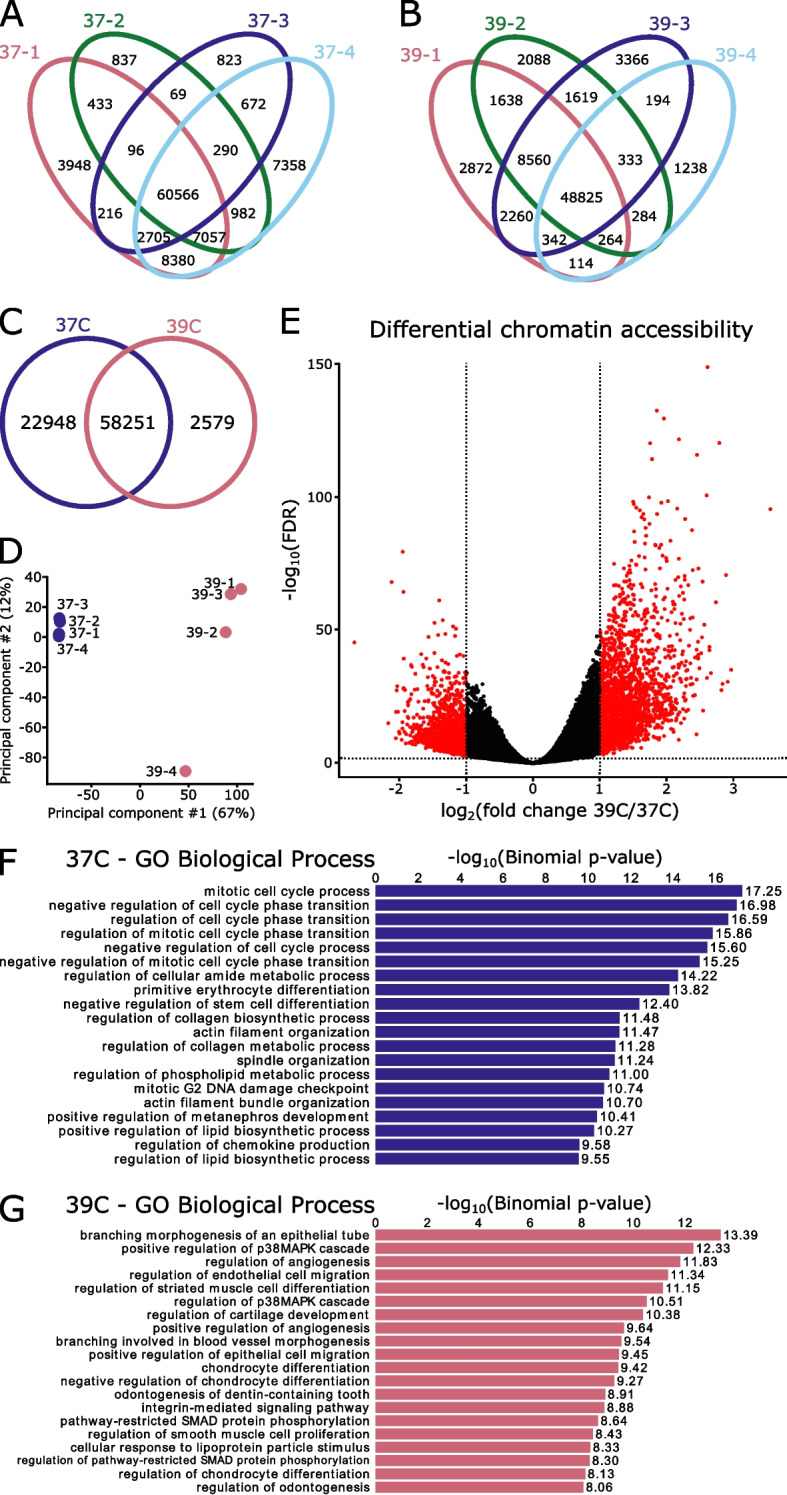
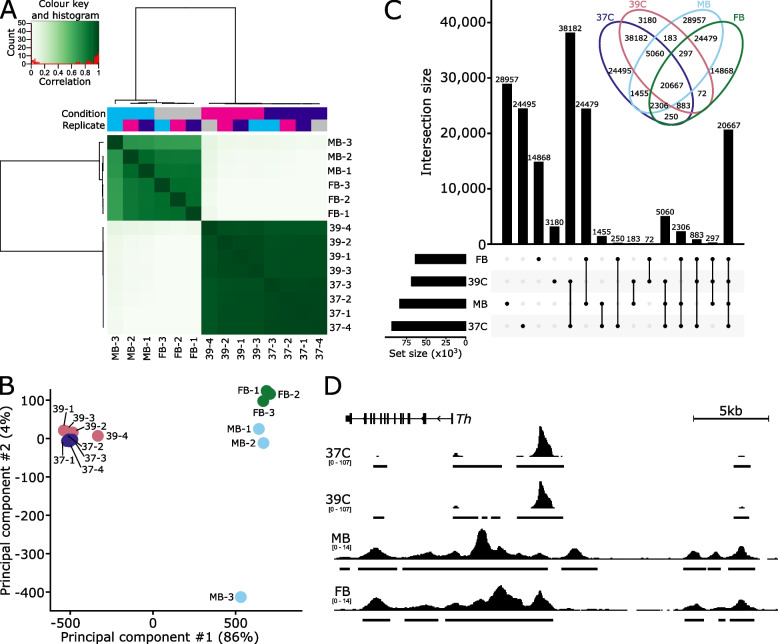
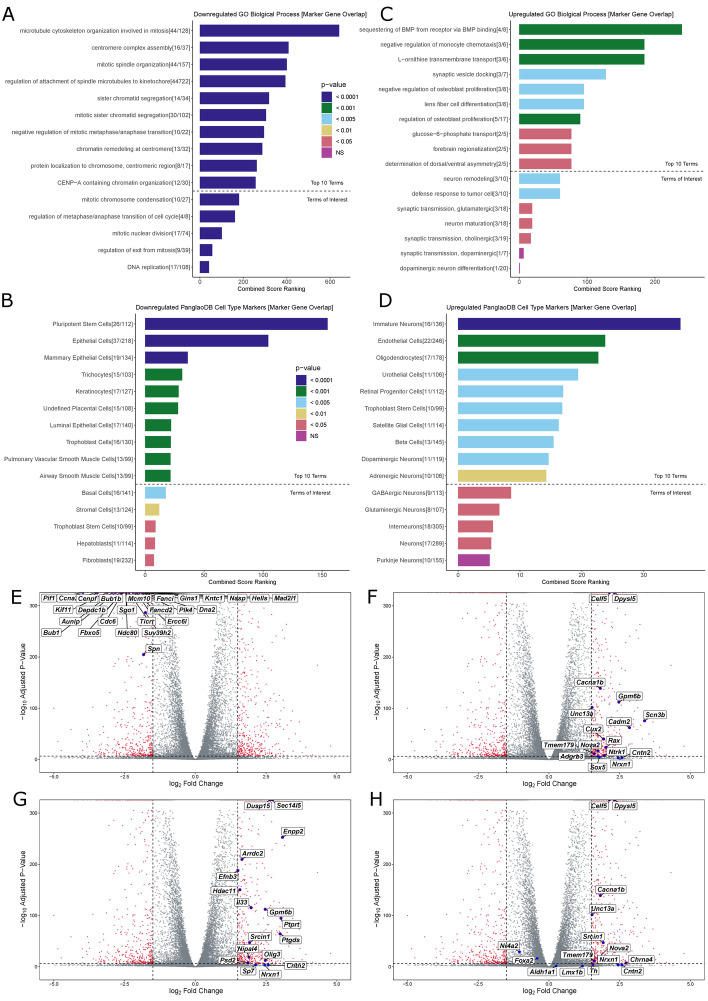
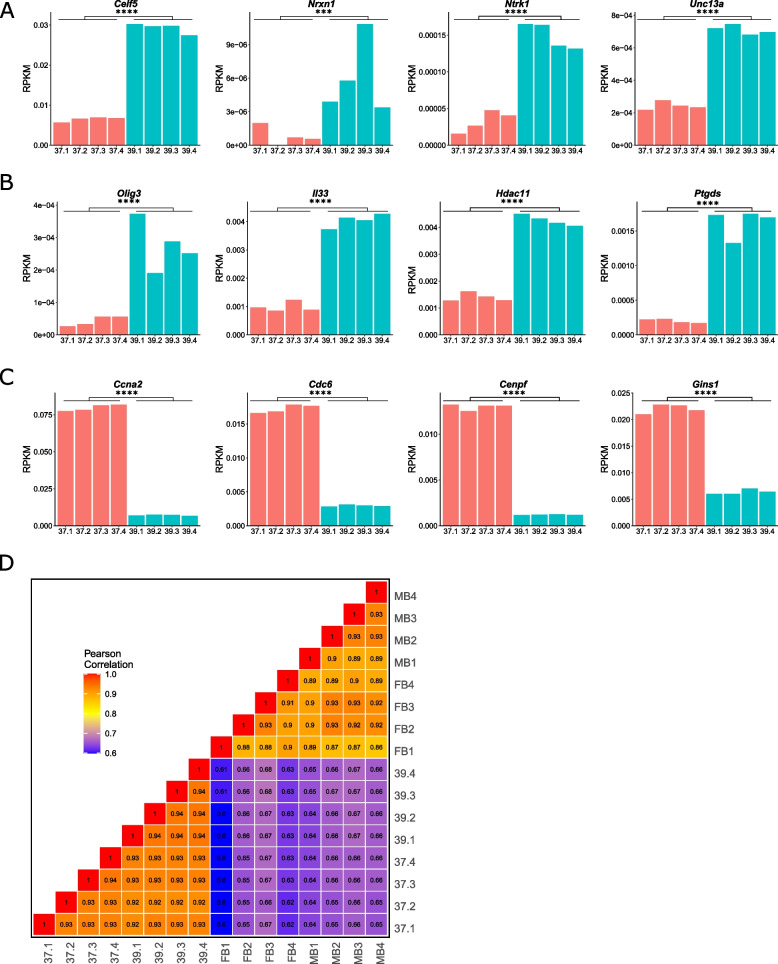
To overcome the ethical and technical limitations of in vivo human disease models, the broader scientific community frequently employs model organism-derived cell lines to investigate disease mechanisms, pathways, and therapeutic strategies. Despite the widespread use of certain in vitro models, many still lack contemporary genomic analysis supporting their use as a proxy for the affected human cells and tissues. Consequently, it is imperative to determine how accurately and effectively any proposed biological surrogate may reflect the biological processes it is assumed to model. One such cellular surrogate of human disease is the established mouse neural precursor cell line, SN4741, which has been used to elucidate mechanisms of neurotoxicity in Parkinson disease for over 25 years. Here, we are using a combination of classic and contemporary genomic techniques - karyotyping, RT-qPCR, single cell RNA-seq, bulk RNA-seq, and ATAC-seq - to characterize the transcriptional landscape, chromatin landscape, and genomic architecture of this cell line, and evaluate its suitability as a proxy for midbrain dopaminergic neurons in the study of Parkinson disease. We find that SN4741 cells possess an unstable triploidy and consistently exhibits low expression of dopaminergic neuron markers across assays, even when the cell line is shifted to the non-permissive temperature that drives differentiation. The transcriptional signatures of SN4741 cells suggest that they are maintained in an undifferentiated state at the permissive temperature and differentiate into immature neurons at the non-permissive temperature; however, they may not be dopaminergic neuron precursors, as previously suggested. Additionally, the chromatin landscapes of SN4741 cells, in both the differentiated and undifferentiated states, are not concordant with the open chromatin profiles of ex vivo, mouse E15.5 forebrain- or midbrain-derived dopaminergic neurons. Overall, our data suggest that SN4741 cells may reflect early aspects of neuronal differentiation but are likely not a suitable proxy for dopaminergic neurons as previously thought. The implications of this study extend broadly, illuminating the need for robust biological and genomic rationale underpinning the use of in vitro models of molecular processes.
Keywords: ATAC-seq; Chromatin accessibility; Disease-relevant model systems; Genomic characterization; Immortalized cell lines; Mouse-derived cell lines; Parkinson disease; RNA-seq; scRNA-seq.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Evaluating the mouse neural precursor line, SN4741, as a suitable proxy for midbrain dopaminergic neurons.bioRxiv [Preprint]. 2023 Feb 1:2023.01.23.525270. doi: 10.1101/2023.01.23.525270. bioRxiv. 2023. Update in: BMC Genomics. 2023 Jun 7;24(1):306. doi: 10.1186/s12864-023-09398-y. PMID: 36747739 Free PMC article. Updated. Preprint.
-
Evaluating the mouse neural precursor line, SN4741, as a suitable proxy for midbrain dopaminergic neurons.Res Sq [Preprint]. 2023 Feb 16:rs.3.rs-2520557. doi: 10.21203/rs.3.rs-2520557/v1. Res Sq. 2023. Update in: BMC Genomics. 2023 Jun 7;24(1):306. doi: 10.1186/s12864-023-09398-y. PMID: 36824793 Free PMC article. Updated. Preprint.
References
-
- Barbosa DJ, Capela JP, de Lourdes BM, Carvalho F. In vitro models for neurotoxicology research. Toxicol Res. 2015;4(4):801–842.
-
- Fisher S, Grice EA, Vinton RM, Bessling SL, Urasaki A, Kawakami K, et al. Evaluating the biological relevance of putative enhancers using Tol2 transposon-mediated transgenesis in zebrafish. Nat Protoc. 2006;1(3):1297–1305. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
