

ALA reverses ABA-induced stomatal closure by modulating PP2AC and SnRK2.6 activity in apple leaves
- PMID: 37287446
- PMCID: PMC10243991
- DOI: 10.1093/hr/uhad067
ALA reverses ABA-induced stomatal closure by modulating PP2AC and SnRK2.6 activity in apple leaves
Abstract
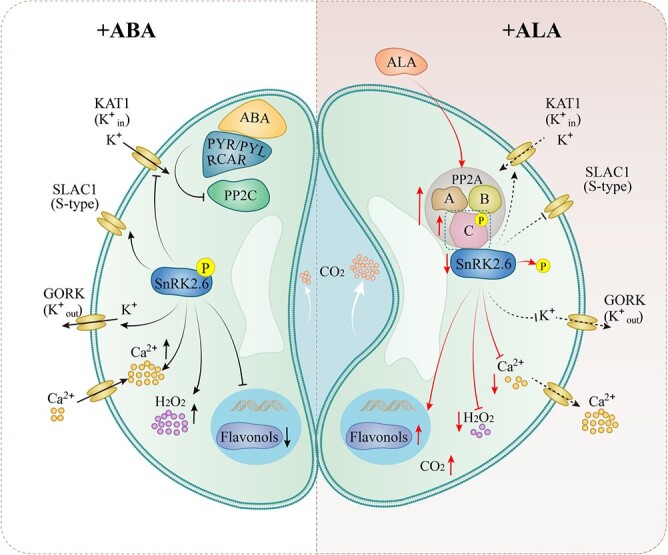
5-Aminolevulinic acid (ALA), known as a new natural plant growth regulator, can reverse abscisic acid (ABA)-induced stomatal closure. The protein phosphatase 2A (PP2A) played an important role in regulation of stomatal movement by ALA and ABA; however, the underlying molecular mechanisms remain unclear. Here, we report that ALA promotes MdPP2A activity and gene expression in the leaf epidermis of apple (Malus × domestica Borkh.), and expression of the catalytic subunit MdPP2AC was most significantly correlated with stomatal aperture. Western blotting showed that ALA enhanced MdPP2AC protein abundance and phosphorylation. Y2H (yeast two hybrid), FLC (firefly luciferase complementation imaging) and BiFC (Bimolecular fluorescence complementation) assays showed that MdPP2AC interacted with several other MdPP2A subunits as well as MdSnRK2.6 (Sucrose non-fermenting 1-related protein kinase 2.6), and the latter interaction was further verified by pull-down and MST (microscale thermophoresis) assays. ALA downregulated ABA-induced MdSnRK2.6 gene expression, kinase activity, and protein phosphorylation. In transiently transgenic apple leaves, OE-MdPP2AC promoted stomatal aperture by reducing Ca2+ and H2O2 levels but increasing flavonol levels in guard cells. Conversely, OE-MdSnRK2.6 induced stomatal closure by increasing Ca2+ and H2O2 but reducing flavonols. Partial silencing of these genes had opposite effects on Ca2+, H2O2, flavonols, and stomatal movement. Application of exogenous ALA stimulated PP2A activity, which promoted SnRK2.6 dephosphorylation and lower kinase activity in wild-type and transgenic apple leaves. We therefore propose that PP2AC, which dephosphorylates SnRK2.6 and represses its enzyme activity, mediates ALA signaling to inhibit ABA-induced stomatal closure in apple leaves.
© The Author(s) 2023. Published by Oxford University Press.
Conflict of interest statement
None declared.
Figures









References
-
- Beale SI. Biosynthesis of 5-aminolevulinic acid. In: Grimm B, Porra RJ, Rüdiger W, Scheer H, eds. Chlorophylls and Bacteriochlorophylls: Biochemistry, Biophysics, Functions and Applications. Springer: The Netherlands, 2006,147–58.
-
- Wang LJ, Jiang WB, Huang BJ. Promotion of 5-aminolevulinic acid on photosynthesis of melon (Cucumis melo) seedlings under low light and chilling stress conditions. Physiol Plant. 2004;121:258–64. - PubMed
-
- Wang LJ, Jiang WB, Liu Het al. . Promotion by 5-aminolevulinic acid of germination of pakchoi(Brassica campestris ssp. Chinensis var. communis Tsen et Lee)seeds under salt stress. J Integr Plant Biol. 2005;47:1084–91.
-
- Wang LJ, Jiang WB, Zhang Zet al. . Biosynthesis and physiological activities of 5-aminolevulinic acid (ALA) and its potential application in agriculture. Plant Physiol Commun. 2003;25:185–92.
-
- Hotta Y, Tanaka T, Takaoka Het al. . New physiological effects of 5-aminolevulinic acid in plants: the increase of photosynthesis, chlorophyll content, and plant growth. Biosci Biotechnol Biochem. 1997;61:2025–8. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
