

Host-specific sensing of coronaviruses and picornaviruses by the CARD8 inflammasome
- PMID: 37289745
- PMCID: PMC10249858
- DOI: 10.1371/journal.pbio.3002144
Host-specific sensing of coronaviruses and picornaviruses by the CARD8 inflammasome
Abstract
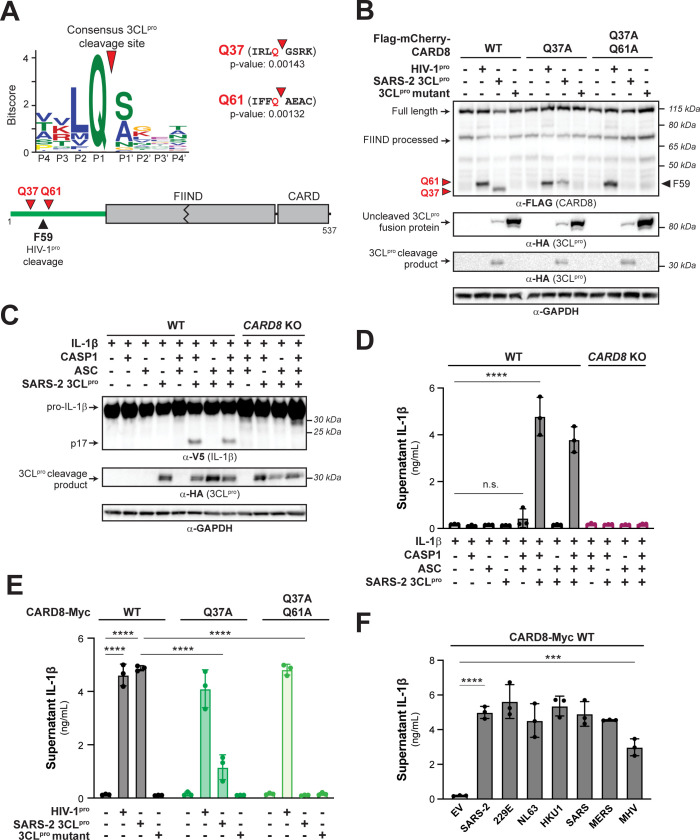
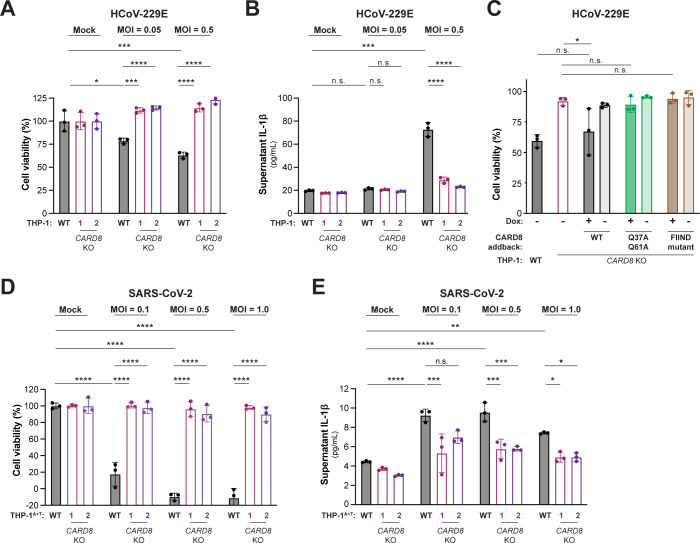
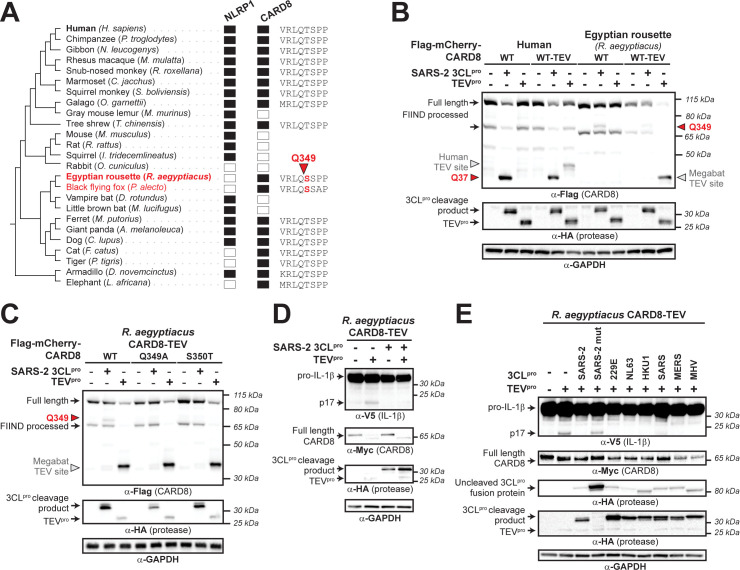
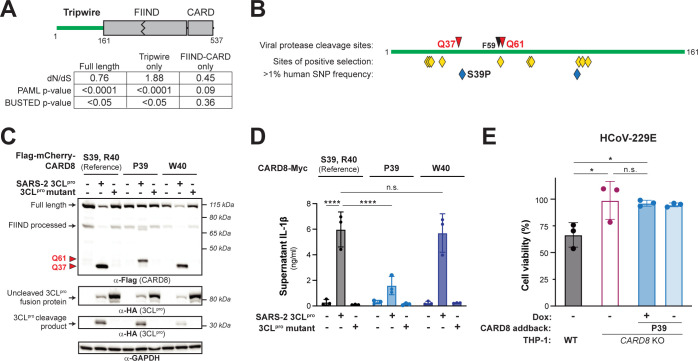
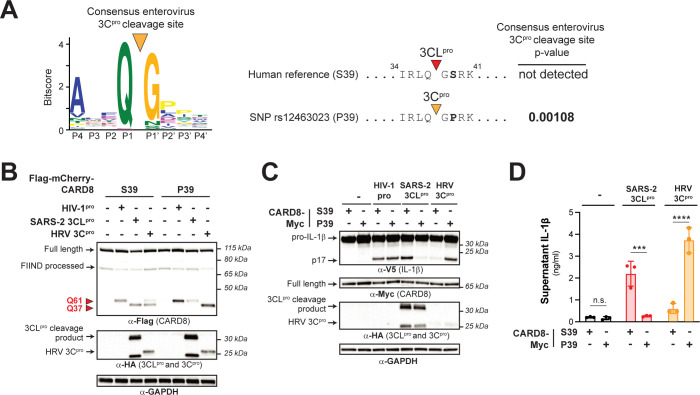
Hosts have evolved diverse strategies to respond to microbial infections, including the detection of pathogen-encoded proteases by inflammasome-forming sensors such as NLRP1 and CARD8. Here, we find that the 3CL protease (3CLpro) encoded by diverse coronaviruses, including Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2), cleaves a rapidly evolving region of human CARD8 and activates a robust inflammasome response. CARD8 is required for cell death and the release of pro-inflammatory cytokines during SARS-CoV-2 infection. We further find that natural variation alters CARD8 sensing of 3CLpro, including 3CLpro-mediated antagonism rather than activation of megabat CARD8. Likewise, we find that a single nucleotide polymorphism (SNP) in humans reduces CARD8's ability to sense coronavirus 3CLpros and, instead, enables sensing of 3C proteases (3Cpro) from select picornaviruses. Our findings demonstrate that CARD8 is a broad sensor of viral protease activities and suggests that CARD8 diversity contributes to inter- and intraspecies variation in inflammasome-mediated viral sensing and immunopathology.
Copyright: © 2023 Tsu et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Update of
-
Host specific sensing of coronaviruses and picornaviruses by the CARD8 inflammasome.bioRxiv [Preprint]. 2022 Sep 22:2022.09.21.508960. doi: 10.1101/2022.09.21.508960. bioRxiv. 2022. Update in: PLoS Biol. 2023 Jun 8;21(6):e3002144. doi: 10.1371/journal.pbio.3002144. PMID: 36172130 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
