

Nitric oxide delivery and heme-assisted S-nitrosation by the bedbug nitrophorin
- PMID: 37290359
- PMCID: PMC10332259
- DOI: 10.1016/j.jinorgbio.2023.112263
Nitric oxide delivery and heme-assisted S-nitrosation by the bedbug nitrophorin
Abstract
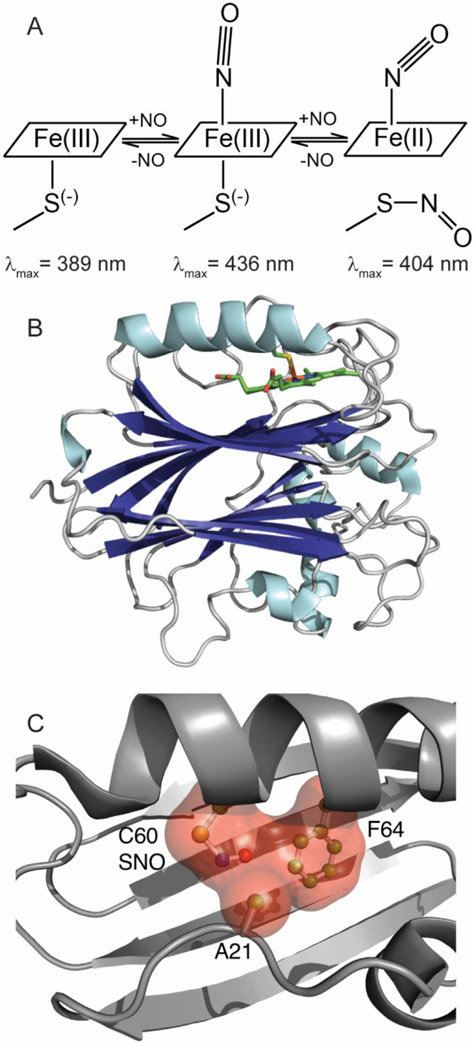
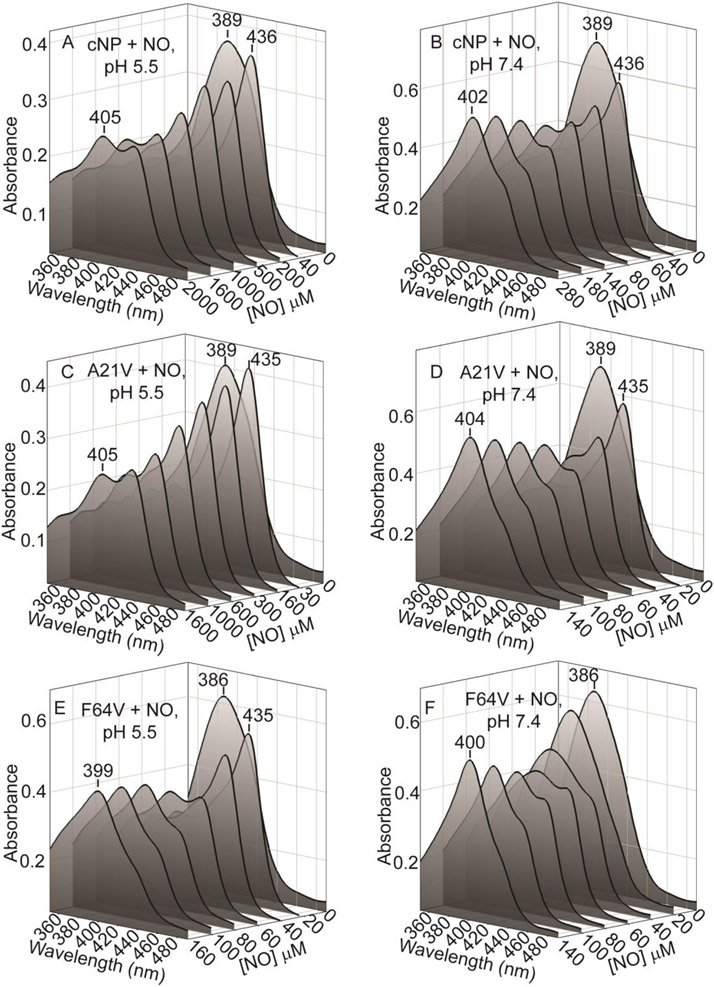
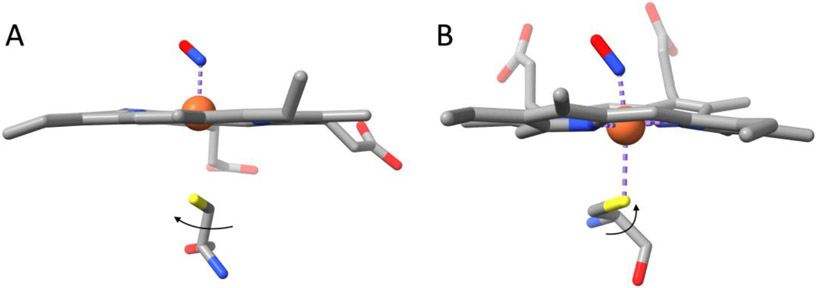
Nitrophorins are heme proteins used by blood feeding insects to deliver nitric oxide (NO) to a victim, leading to vasodilation and antiplatelet activity. Cimex lectularius (bedbug) nitrophorin (cNP) accomplishes this with a cysteine ligated ferric (Fe(III)) heme. In the acidic environment of the insect's salivary glands, NO binds tightly to cNP. During a blood meal, cNP-NO is delivered to the feeding site where dilution and increased pH lead to NO release. In a previous study, cNP was shown to not only bind heme, but to also nitrosate the proximal cysteine, leading to Cys-NO (SNO) formation. SNO formation requires oxidation of the proximal cysteine, which was proposed to be metal-assisted through accompanying reduction of ferric heme and formation of Fe(II)-NO. Here, we report the 1.6 Å crystal structure of cNP first chemically reduced and then exposed to NO, and show that Fe(II)-NO is formed but SNO is not, supporting a metal-assisted SNO formation mechanism. Crystallographic and spectroscopic studies of mutated cNP show that steric crowding of the proximal site inhibits SNO formation while a sterically relaxed proximal site enhances SNO formation, providing insight into specificity for this poorly understood modification. Experiments examining the pH dependence for NO implicate direct protonation of the proximal cysteine as the underlying mechanism. At lower pH, thiol heme ligation predominates, leading to a smaller trans effect and 60-fold enhanced NO affinity (Kd = 70 nM). Unexpectedly, we find that thiol formation interferes with SNO formation, suggesting cNP-SNO is unlikely to form in the insect salivary glands.
Keywords: Heme protein; Nitric oxide; Nitrophorin; Nitrosylation; S-nitrosocysteine.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures





References
-
- Walker FA. Nitric oxide interaction with insect nitrophorins and thoughts on the electron configuration of the {FeNO}6 complex. J. Inorg. Biochem 2005, 99:216–236. - PubMed
-
- Montfort WR, Weichsel A, Andersen JF. Nitrophorins and related antihemostatic lipocalins from Rhodnius prolixus and other blood-sucking arthropods. Biochim. Biophys. Acta 2000, 1482:110–118. - PubMed
-
- Maes EM, Roberts SA, Weichsel A, Montfort WR. Ultrahigh Resolution Structures of Nitrophorin 4: Heme Distortion in Ferrous CO and NO Complexes. Biochemistry. 2005, 44:12690–12699. - PubMed
-
- Kondrashov DA, Roberts SA, Weichsel A, Montfort WR. Protein functional cycle viewed at atomic resolution: conformational change and mobility in nitrophorin 4 as a function of pH and NO binding. Biochemistry. 2004, 43:13637–13647. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
