

This is a preprint.
A new antibiotic from an uncultured bacterium binds to an immutable target
- PMID: 37292624
- PMCID: PMC10245560
- DOI: 10.1101/2023.05.15.540765
A new antibiotic from an uncultured bacterium binds to an immutable target
Update in
-
An antibiotic from an uncultured bacterium binds to an immutable target.Cell. 2023 Sep 14;186(19):4059-4073.e27. doi: 10.1016/j.cell.2023.07.038. Epub 2023 Aug 22. Cell. 2023. PMID: 37611581
Abstract
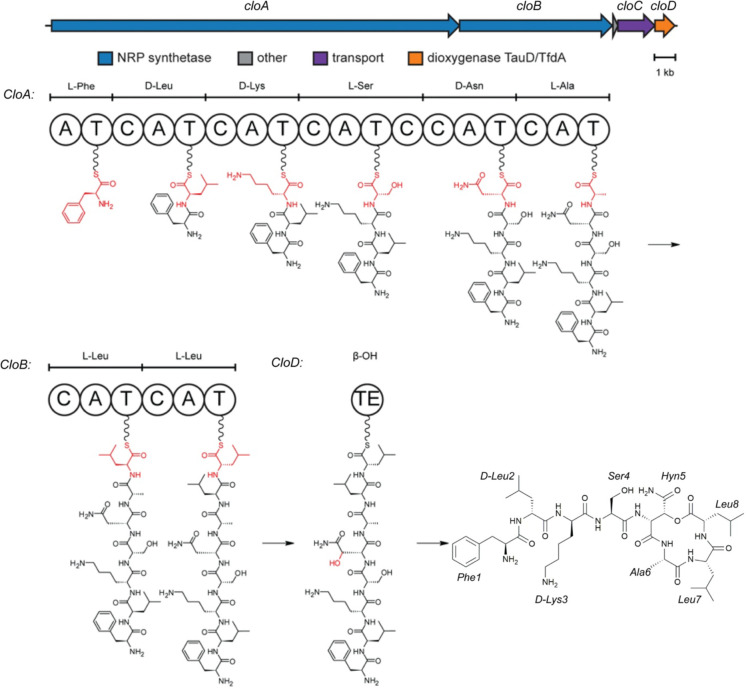
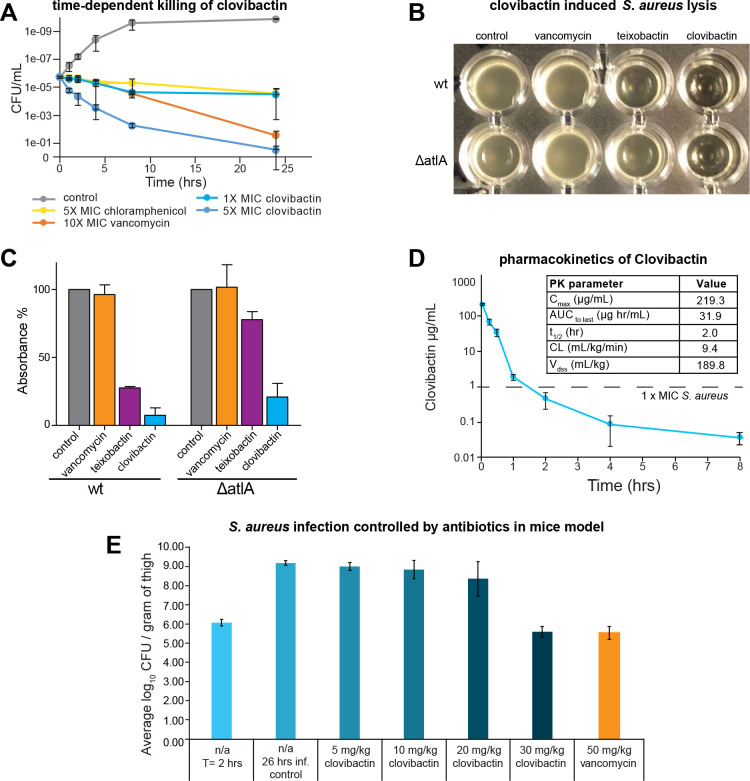
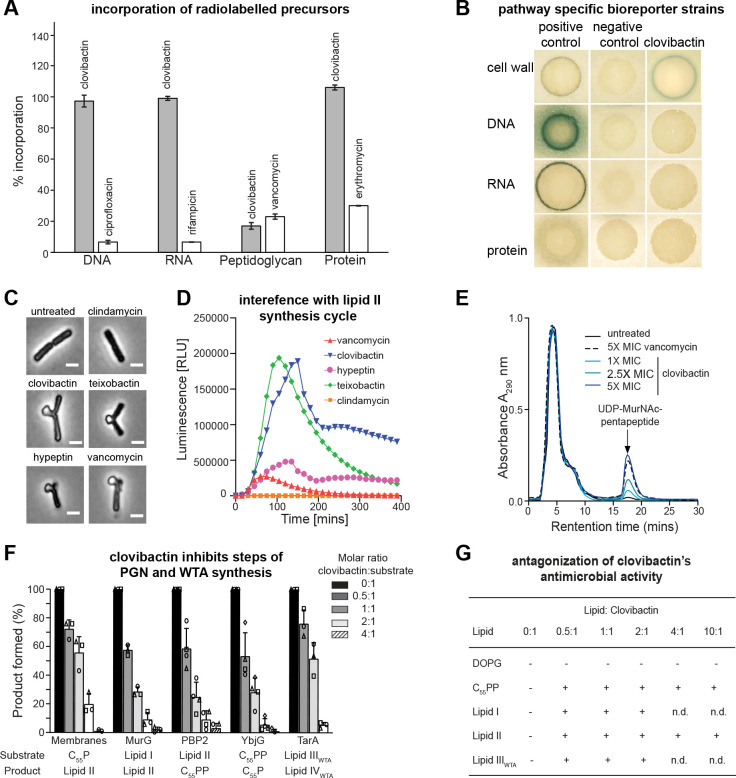
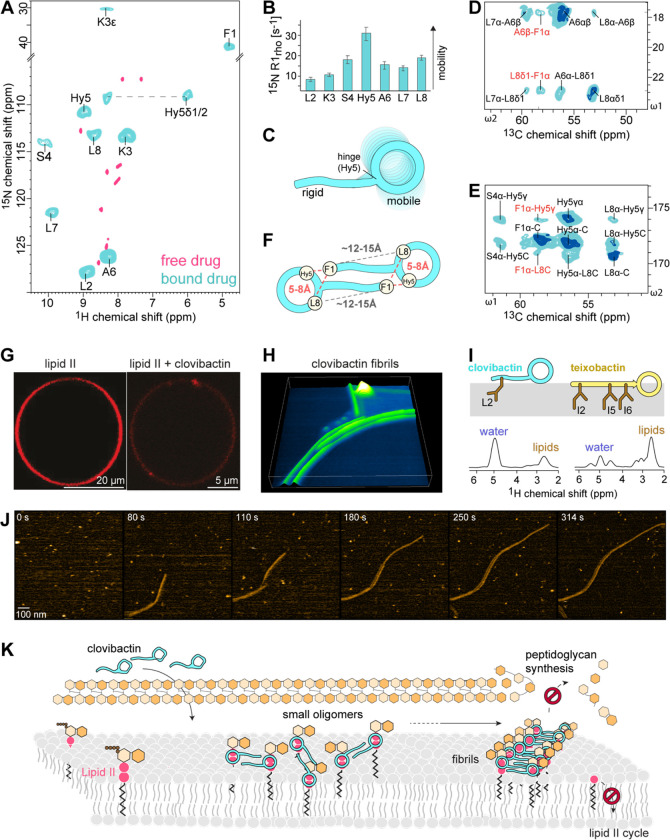
Antimicrobial resistance is a leading mortality factor worldwide. Here we report the discovery of clovibactin, a new antibiotic, isolated from uncultured soil bacteria. Clovibactin efficiently kills drug-resistant bacterial pathogens without detectable resistance. Using biochemical assays, solid-state NMR, and atomic force microscopy, we dissect its mode of action. Clovibactin blocks cell wall synthesis by targeting pyrophosphate of multiple essential peptidoglycan precursors (C 55 PP, Lipid II, Lipid WTA ). Clovibactin uses an unusual hydrophobic interface to tightly wrap around pyrophosphate, but bypasses the variable structural elements of precursors, accounting for the lack of resistance. Selective and efficient target binding is achieved by the irreversible sequestration of precursors into supramolecular fibrils that only form on bacterial membranes that contain lipid-anchored pyrophosphate groups. Uncultured bacteria offer a rich reservoir of antibiotics with new mechanisms of action that could replenish the antimicrobial discovery pipeline.
Conflict of interest statement
Competing Interests
The following authors, A. J. Peoples, C. Achorn, A. Nitti, A. L. Spoering, L. L. Ling, D. E. Hughes, and K. Lewis, declare competing financial interests as they are employees and consultants of NovoBiotic Pharmaceuticals. A patent US 11,203,616 B2 was issued 12/21/2021 and describes the use of clovibactin (Novo29) and as an antibiotic, as well as the pharmaceutical composition and antibiotic use of derivatives. The other authors have no competing interests.
Figures

References
-
- Baldus M., and Meier B.H. (1996). Total Correlation Spectroscopy in the Solid State. The Use of Scalar Couplings to Determine the Through-Bond Connectivity. Journal of Magnetic Resonance, Series A 121, 65–69.
-
- Breukink E., van Heusden H.E., Vollmerhaus P.J., Swiezewska E., Brunner L., Walker S., Heck A.J.R., and de Kruijff B. (2003). Lipid II is an intrinsic component of the pore induced by nisin in bacterial membranes. J Biol Chem 278, 19898–19903. - PubMed
-
- Brown E.D., and Wright G.D. (2016). Antibacterial drug discovery in the resistance era. Nature 529, 336–343. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous