

This is a preprint.
Lifetime of ground conformational state determines the activity of structured RNA
- PMID: 37292668
- PMCID: PMC10246243
- DOI: 10.21203/rs.3.rs-2879957/v1
Lifetime of ground conformational state determines the activity of structured RNA
Update in
-
Lifetime of ground conformational state determines the activity of structured RNA.Nat Chem Biol. 2025 Jul;21(7):1021-1029. doi: 10.1038/s41589-025-01843-1. Epub 2025 Feb 12. Nat Chem Biol. 2025. PMID: 39939412 Free PMC article.
Abstract
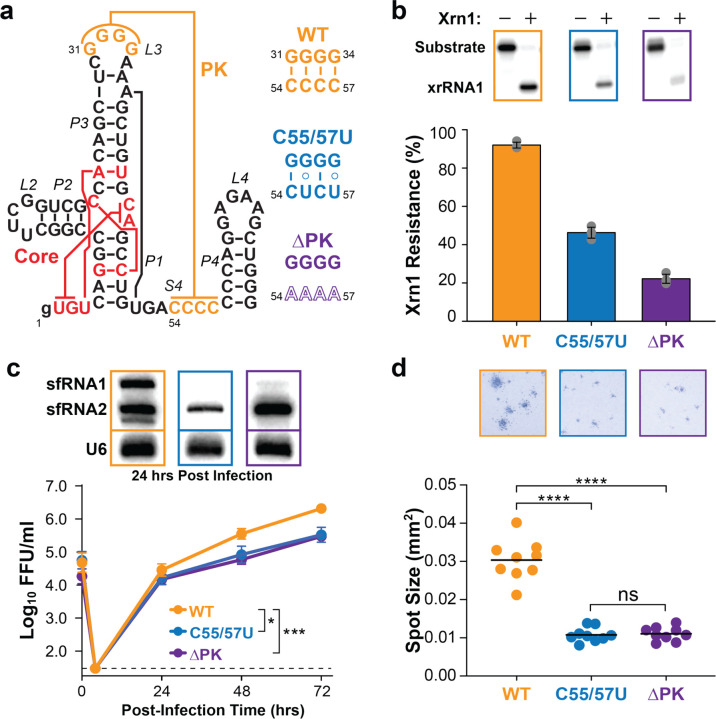
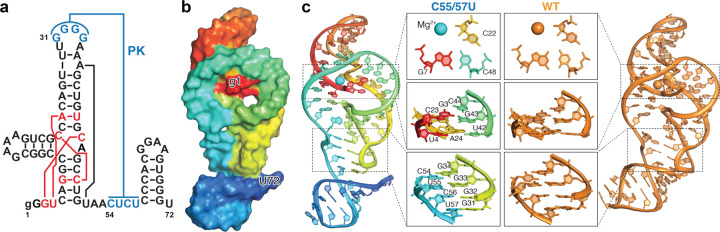
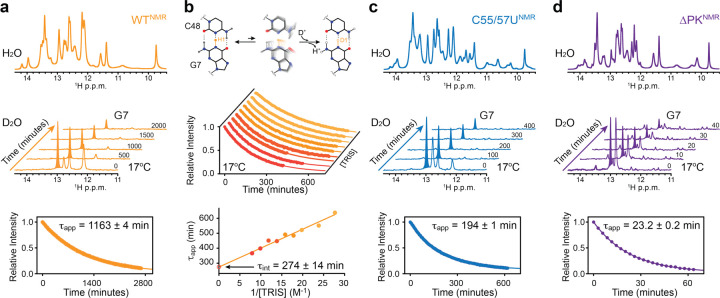
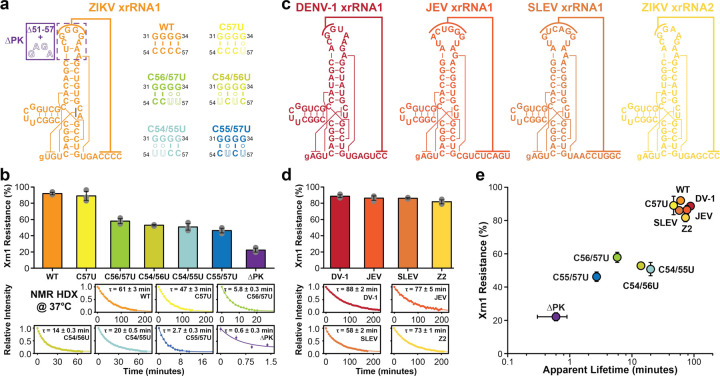
Biomolecules continually sample alternative conformations. Consequently, even the most energetically favored ground conformational state has a finite lifetime. Here, we show that, in addition to the 3D structure, the lifetime of a ground conformational state determines its biological activity. Using hydrogen-deuterium exchange nuclear magnetic resonance spectroscopy, we found that Zika virus exoribonuclease-resistant RNA (xrRNA) encodes a ground conformational state with a lifetime that is ~105-107 longer than that of canonical base pairs. Mutations that shorten the apparent lifetime of the ground state without affecting its 3D structure decreased exoribonuclease resistance in vitro and impaired virus replication in cells. Additionally, we observed this exceptionally long-lived ground state in xrRNAs from diverse infectious mosquito-borne flaviviruses. These results demonstrate the biological significance of the lifetime of a preorganized ground state and further suggest that elucidating the lifetimes of dominant 3D structures of biomolecules may be crucial for understanding their behaviors and functions.
Figures

References
-
- Henzler-Wildman K. & Kern D. Dynamic personalities of proteins. Nature 450, 964–972 (2007). - PubMed
-
- Alderson T. R. & Kay L. E. NMR spectroscopy captures the essential role of dynamics in regulating biomolecular function. Cell 184, 577–595 (2021). - PubMed
-
- Boehr D. D., McElheny D., Dyson H. J. & Wright P. E. The dynamic energy landscape of dihydrofolate reductase catalysis. Science 313, 1638–1642 (2006). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
