

This is a preprint.
Cytosolic iron-sulfur protein assembly system identifies clients by a C-terminal tripeptide
- PMID: 37292740
- PMCID: PMC10245660
- DOI: 10.1101/2023.05.19.541488
Cytosolic iron-sulfur protein assembly system identifies clients by a C-terminal tripeptide
Update in
-
Cytosolic iron-sulfur protein assembly system identifies clients by a C-terminal tripeptide.Proc Natl Acad Sci U S A. 2023 Oct 31;120(44):e2311057120. doi: 10.1073/pnas.2311057120. Epub 2023 Oct 26. Proc Natl Acad Sci U S A. 2023. PMID: 37883440 Free PMC article.
Abstract
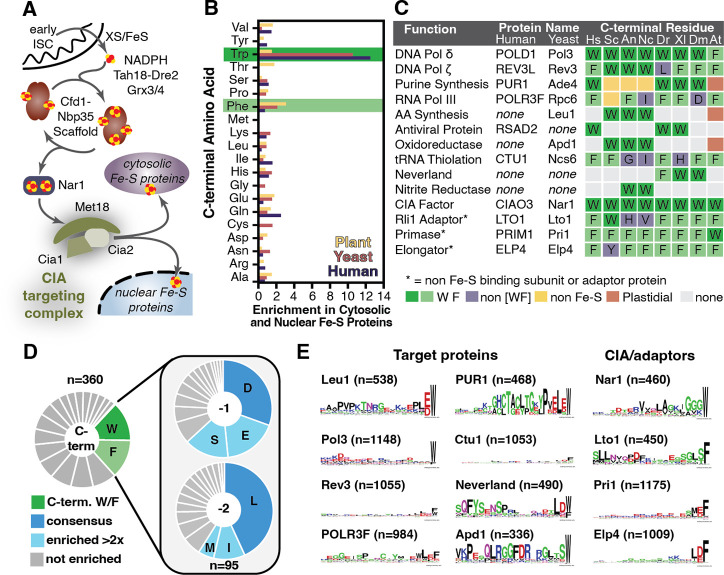
The eukaryotic cytosolic Fe-S protein assembly (CIA) machinery inserts iron-sulfur (Fe-S) clusters into cytosolic and nuclear proteins. In the final maturation step, the Fe-S cluster is transferred to the apo-proteins by the CIA-targeting complex (CTC). However, the molecular recognition determinants of client proteins are unknown. We show that a conserved [LIM]-[DES]-[WF]-COO- tripeptide present at the C-terminus of clients is necessary and sufficient for binding to the CTC in vitro and directing Fe-S cluster delivery in vivo. Remarkably, fusion of this TCR (target complex recognition) signal enables engineering of cluster maturation on a non-native protein via recruitment of the CIA machinery. Our study significantly advances our understanding of Fe-S protein maturation and paves the way for bioengineering applications.
Conflict of interest statement
Competing interests: Authors declare that they have no competing interests.
Figures



References
-
- Braymer J. J., Freibert S. A., Rakwalska-Bange M., Lill R., Mechanistic concepts of iron-sulfur protein biogenesis in Biology. Biochim Biophys Acta Mol Cell Res 1868, 118863 (2021). - PubMed
-
- Weon J. L., Yang S. W., Potts P. R., Cytosolic Iron-Sulfur Assembly Is Evolutionarily Tuned by a Cancer-Amplified Ubiquitin Ligase. Mol Cell 69, 113–125.e116 (2018). - PubMed
-
- Honarmand Ebrahimi K. et al., Iron-sulfur clusters as inhibitors and catalysts of viral replication. Nat Chem 14, 253–266 (2022). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous