

This is a preprint.
BAX activation in mouse retinal ganglion cells occurs in two temporally and mechanistically distinct steps
- PMID: 37292963
- PMCID: PMC10246290
- DOI: 10.21203/rs.3.rs-2846437/v1
BAX activation in mouse retinal ganglion cells occurs in two temporally and mechanistically distinct steps
Update in
-
BAX activation in mouse retinal ganglion cells occurs in two temporally and mechanistically distinct steps.Mol Neurodegener. 2023 Sep 26;18(1):67. doi: 10.1186/s13024-023-00659-8. Mol Neurodegener. 2023. PMID: 37752598 Free PMC article.
Abstract
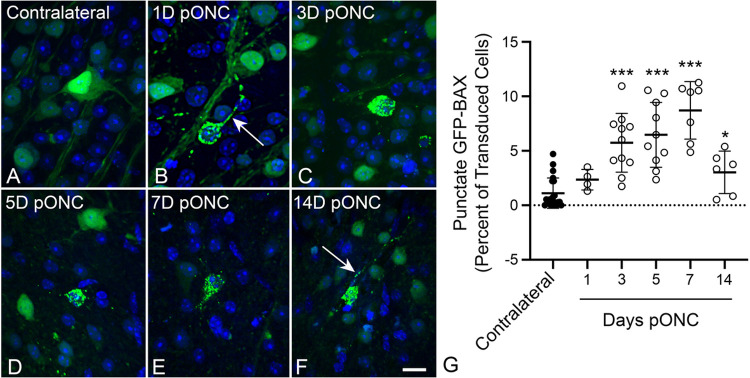
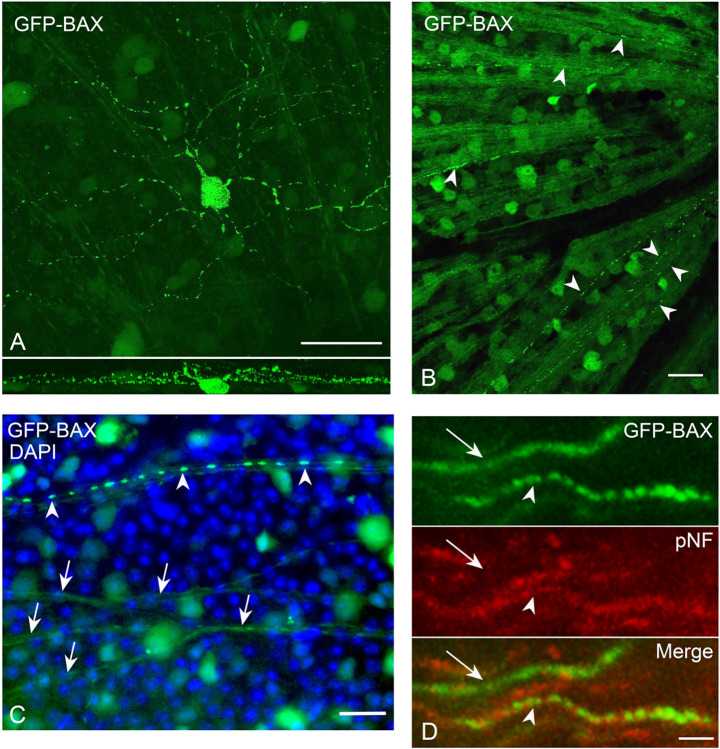
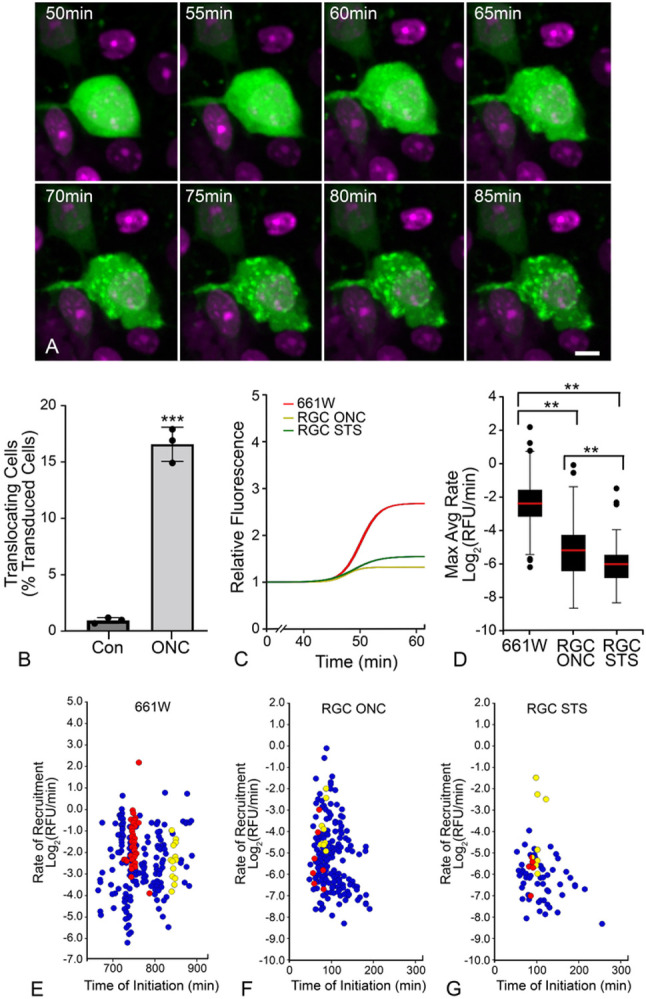
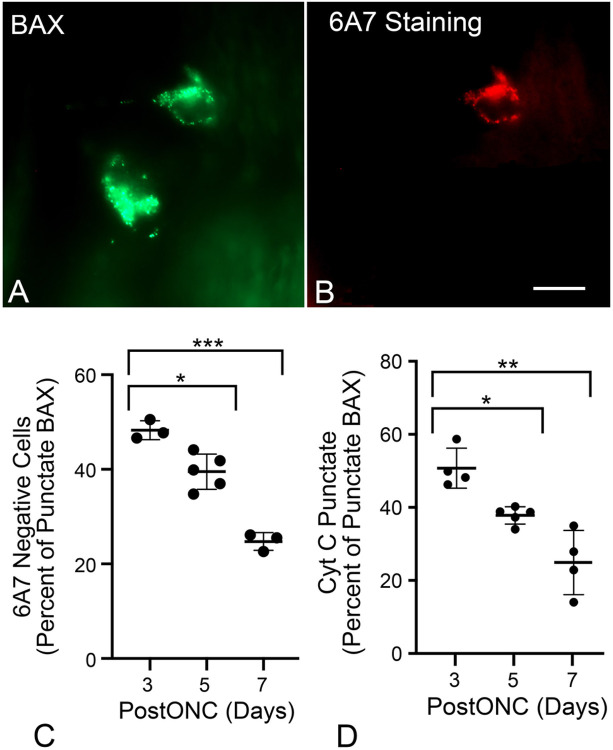
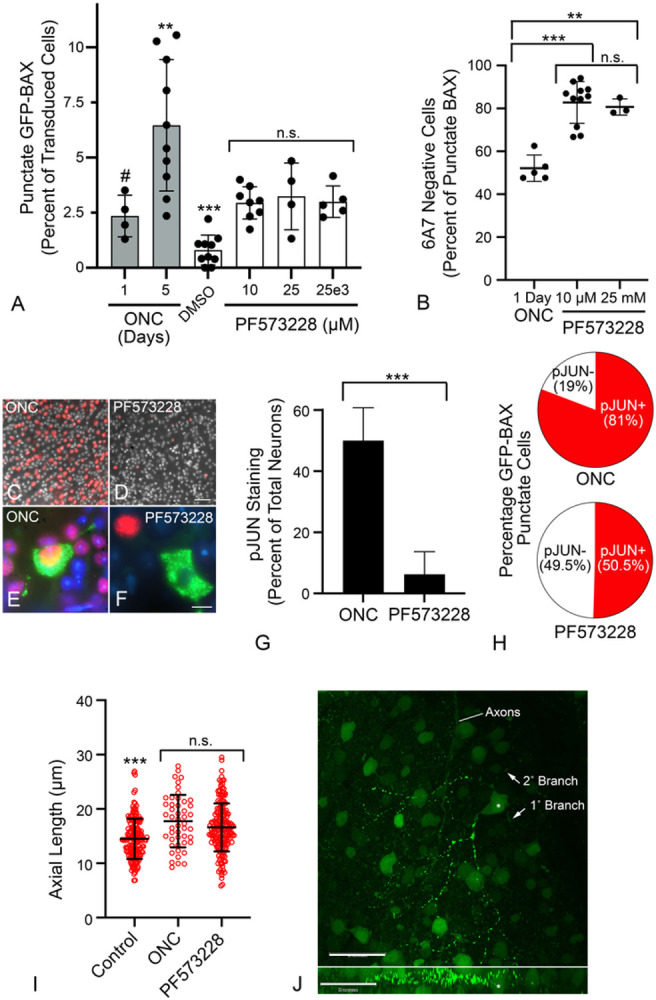
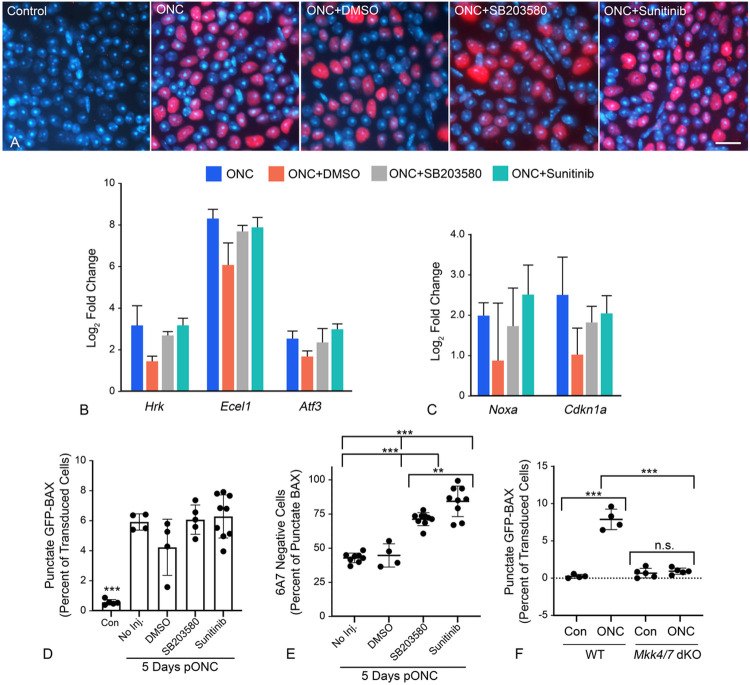
Background Pro-apoptotic BAX is a central mediator of retinal ganglion cell (RGC) death after optic nerve damage. BAX activation occurs in two stages including translocation of latent BAX to the mitochondrial outer membrane (MOM) and then permeabilization of the MOM to facilitate the release of apoptotic signaling molecules. As a critical component of RGC death, BAX is an attractive target for neuroprotective therapies and an understanding of the kinetics of BAX activation and the mechanisms controlling the two stages of this process in RGCs is potentially valuable in informing the development of a neuroprotective strategy. Methods The kinetics of BAX translocation were assessed by both static and live-cell imaging of a GFP-BAX fusion protein introduced into RGCs using AAV2-mediated gene transfer in mice. Activation of BAX was achieved using an acute optic nerve crush (ONC) protocol. Live-cell imaging of GFP-BAX was achieved using explants of mouse retina harvested 7 days after ONC. Kinetics of translocation in RGCs were compared to GFP-BAX translocation in 661W tissue culture cells. Permeabilization of GFP-BAX was assessed by staining with the 6A7 monoclonal antibody, which recognizes a conformational change in this protein after MOM insertion. Assessment of individual kinases associated with both stages of activation was made using small molecule inhibitors injected into the vitreous either independently or in concert with ONC surgery. The contribution of the Dual Leucine Zipper-JUN-N-Terminal Kinase cascade was evaluated using mice with a double conditional knock-out of both Mkk4 and Mkk7 . Results ONC induces the translocation of GFP-BAX in RGCs at a slower rate and with less intracellular synchronicity than 661W cells, but exhibits less variability among mitochondrial foci within a single cell. GFP-BAX was also found to translocate in all compartments of an RGC including the dendritic arbor and axon. Approximately 6% of translocating RGCs exhibited retrotranslocation of BAX immediately following translocation. Unlike tissue culture cells, which exhibit simultaneous translocation and permeabilization, RGCs exhibited a significant delay between these two stages, similar to detached cells undergoing anoikis. Translocation, with minimal permeabilization could be induced in a subset of RGCs using an inhibitor of Focal Adhesion Kinase (PF573228). Permeabilization after ONC, in a majority of RGCs, could be inhibited with a broad spectrum kinase inhibitor (sunitinib) or a selective inhibitor for p38/MAPK14 (SB203580). Intervention of DLK-JNK axis signaling abrogated GFP-BAX translocation after ONC. Conclusions A comparison between BAX activation kinetics in tissue culture cells and in cells of a complex tissue environment shows distinct differences indicating that caution should be used when translating findings from one condition to the other. RGCs exhibit both a delay between translocation and permeabilization and the ability for translocated BAX to be retrotranslocated, suggesting several stages at which intervention of the activation process could be exploited in the design of a therapeutic strategy.
Figures

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
