

This is a preprint.
Overlapping representations of food and social stimuli in VTA dopamine neurons
- PMID: 37293057
- PMCID: PMC10245666
- DOI: 10.1101/2023.05.17.541104
Overlapping representations of food and social stimuli in VTA dopamine neurons
Update in
-
Overlapping representations of food and social stimuli in mouse VTA dopamine neurons.Neuron. 2023 Nov 15;111(22):3541-3553.e8. doi: 10.1016/j.neuron.2023.08.003. Epub 2023 Aug 31. Neuron. 2023. PMID: 37657441 Free PMC article.
Abstract
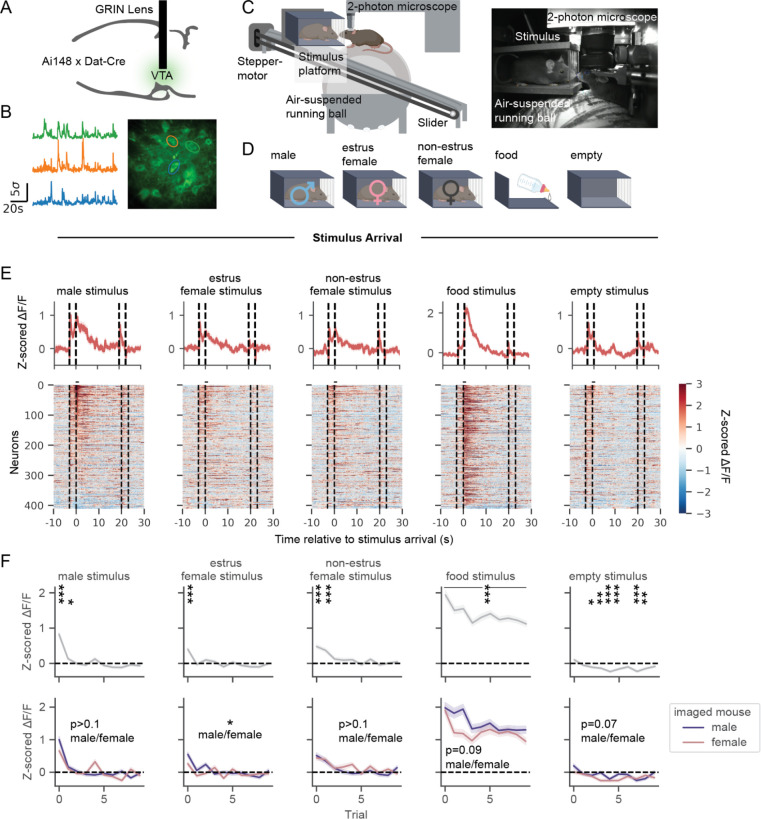
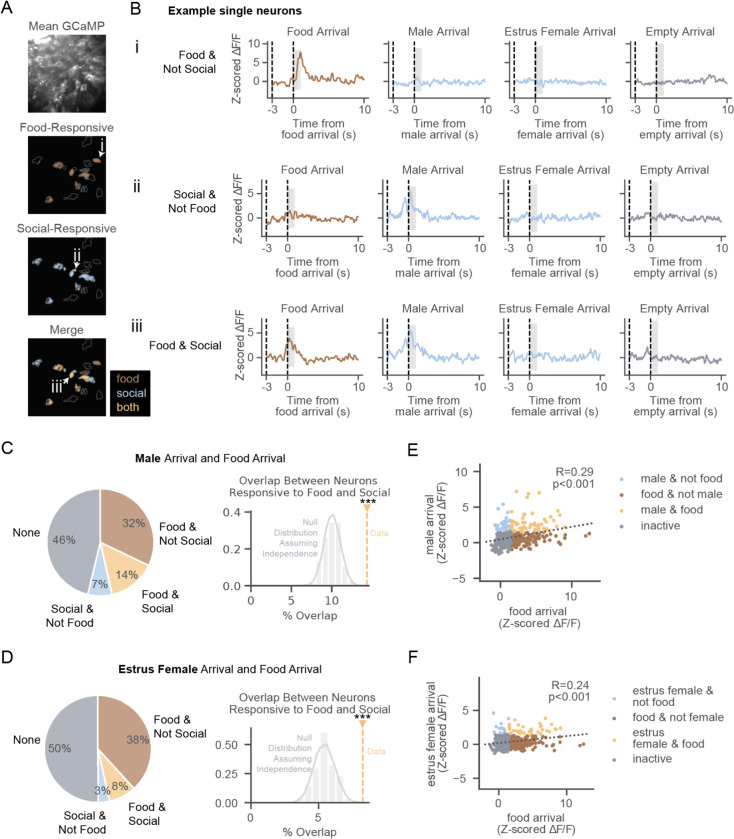
Dopamine neurons of the ventral tegmental area (VTA DA ) respond to food and social stimuli and contribute to both forms of motivation. However, it is unclear if the same or different VTA DA neurons encode these different stimuli. To address this question, we performed 2-photon calcium imaging in mice presented with food and conspecifics, and found statistically significant overlap in the populations responsive to both stimuli. Both hunger and opposite-sex social experience further increased the proportion of neurons that respond to both stimuli, implying that modifying motivation for one stimulus affects responses to both stimuli. In addition, single-nucleus RNA sequencing revealed significant co-expression of feeding- and social-hormone related genes in individual VTA DA neurons. Taken together, our functional and transcriptional data suggest overlapping VTA DA populations underlie food and social motivation.
Figures


References
-
- Abizaid Alfonso, Liu Zhong-Wu, Andrews Zane B., Shanabrough Marya, Borok Erzsebet, Elsworth John D., Roth Robert H., et al. 2006. “Ghrelin Modulates the Activity and Synaptic Input Organization of Midbrain Dopamine Neurons While Promoting Appetite.” The Journal of Clinical Investigation 116 (12): 3229–39. - PMC - PubMed
-
- Adkins-Regan Elizabeth. 2009. “Neuroendocrinology of Social Behavior.” ILAR Journal / National Research Council, Institute of Laboratory Animal Resources 50 (1): 5–14. - PubMed
-
- Adriaenssens Alice E., Biggs Emma K., Darwish Tamana, Tadross John, Sukthankar Tanmay, Girish Milind, Joseph Polex-Wolf, et al. 2019. “Glucose-Dependent Insulinotropic Polypeptide Receptor-Expressing Cells in the Hypothalamus Regulate Food Intake.” Cell Metabolism. 10.1016/j.cmet.2019.07.013. - DOI - PMC - PubMed
-
- Ahima R. S., Prabakaran D., Mantzoros C., Qu D., Lowell B., Maratos-Flier E., and Flier J. S.. 1996. “Role of Leptin in the Neuroendocrine Response to Fasting.” Nature 382 (6588): 250–52. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources