

This is a preprint.
Asymmetric apical domain states of mitochondrial Hsp60 coordinate substrate engagement and chaperonin assembly
- PMID: 37293102
- PMCID: PMC10245740
- DOI: 10.1101/2023.05.15.540872
Asymmetric apical domain states of mitochondrial Hsp60 coordinate substrate engagement and chaperonin assembly
Update in
-
Asymmetric apical domain states of mitochondrial Hsp60 coordinate substrate engagement and chaperonin assembly.Nat Struct Mol Biol. 2024 Dec;31(12):1848-1858. doi: 10.1038/s41594-024-01352-0. Epub 2024 Jul 1. Nat Struct Mol Biol. 2024. PMID: 38951622 Free PMC article.
Abstract
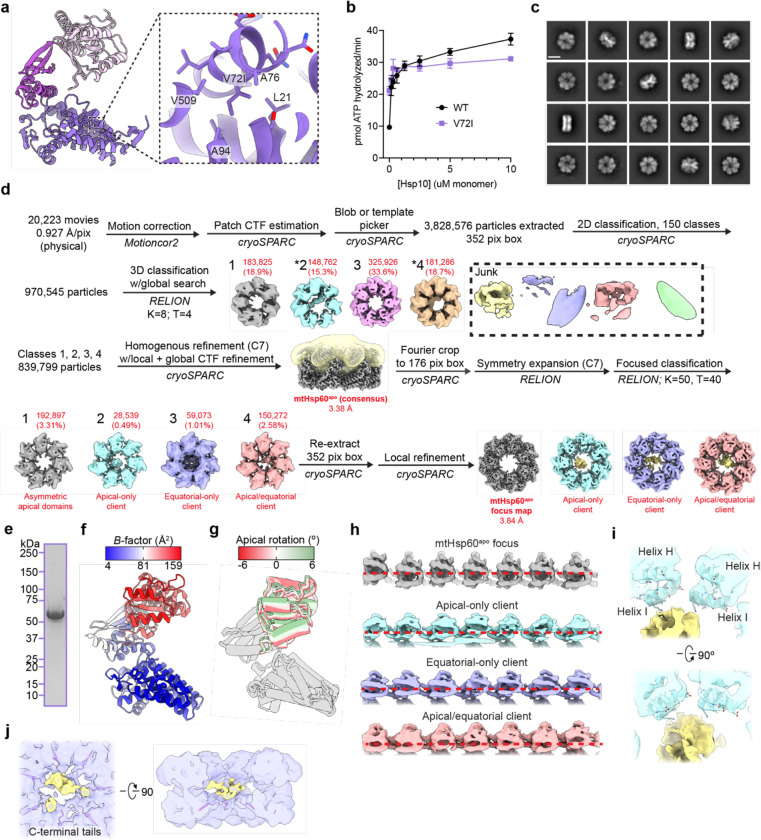
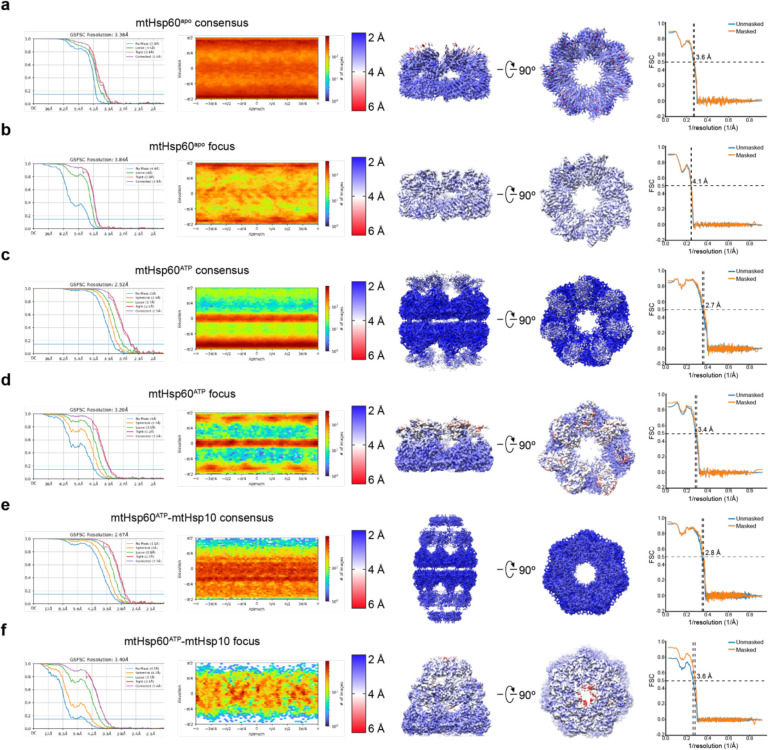
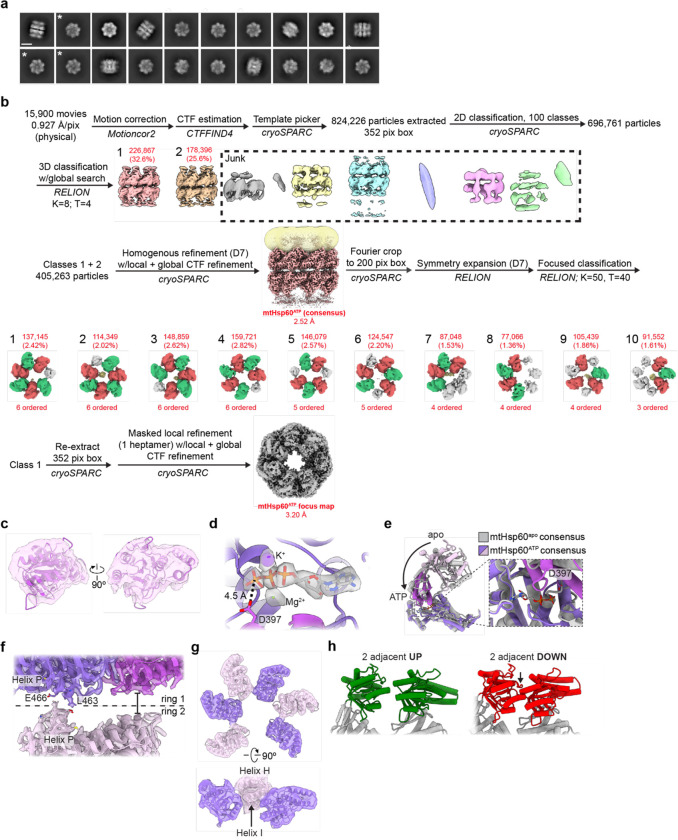
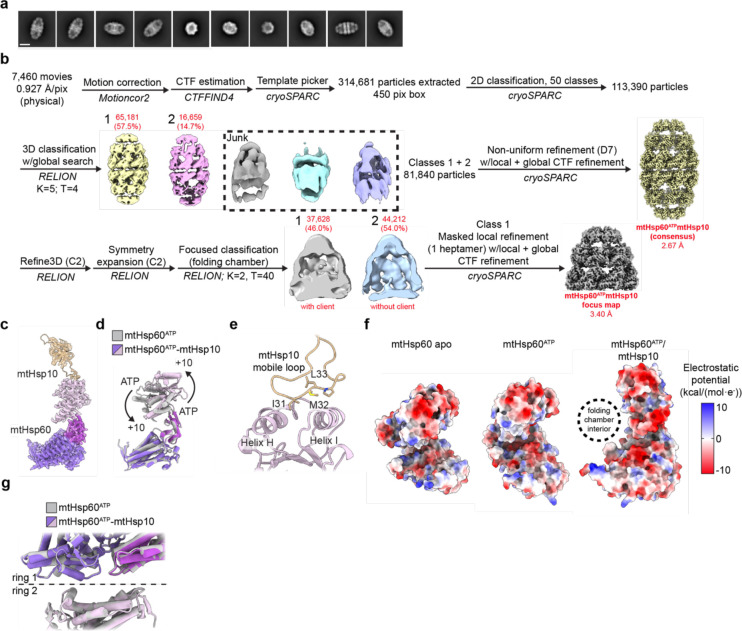
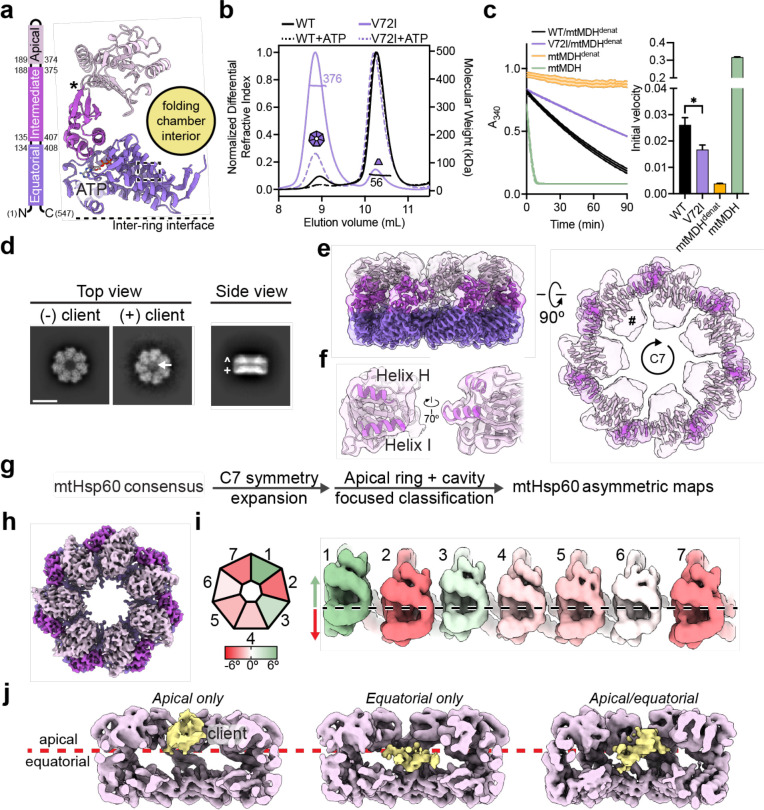
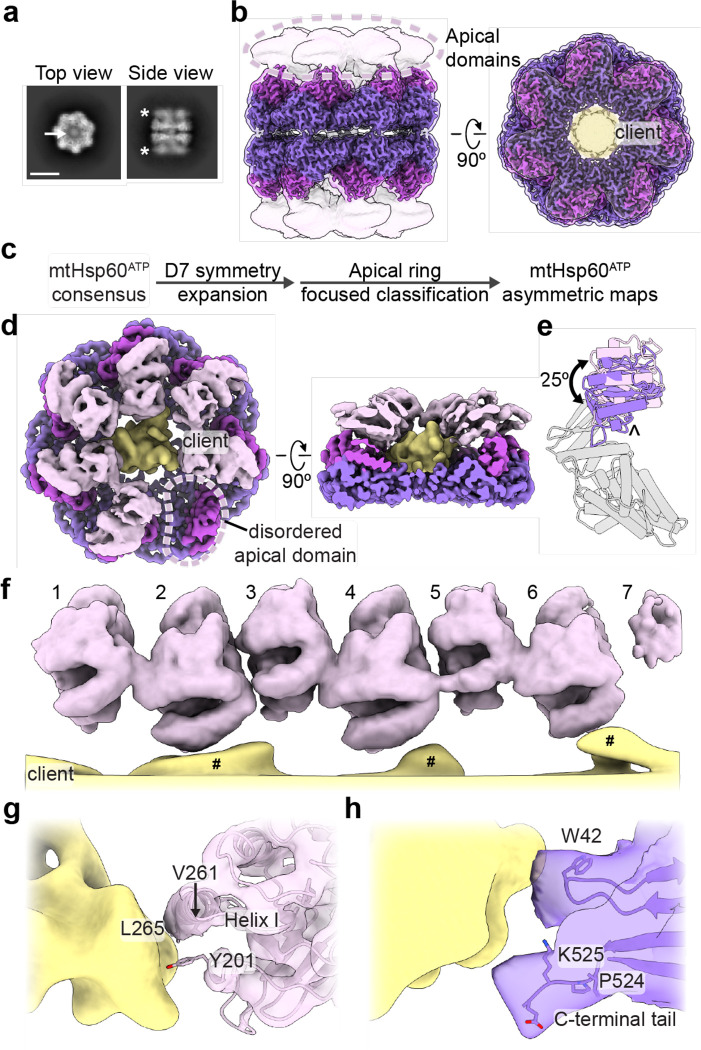
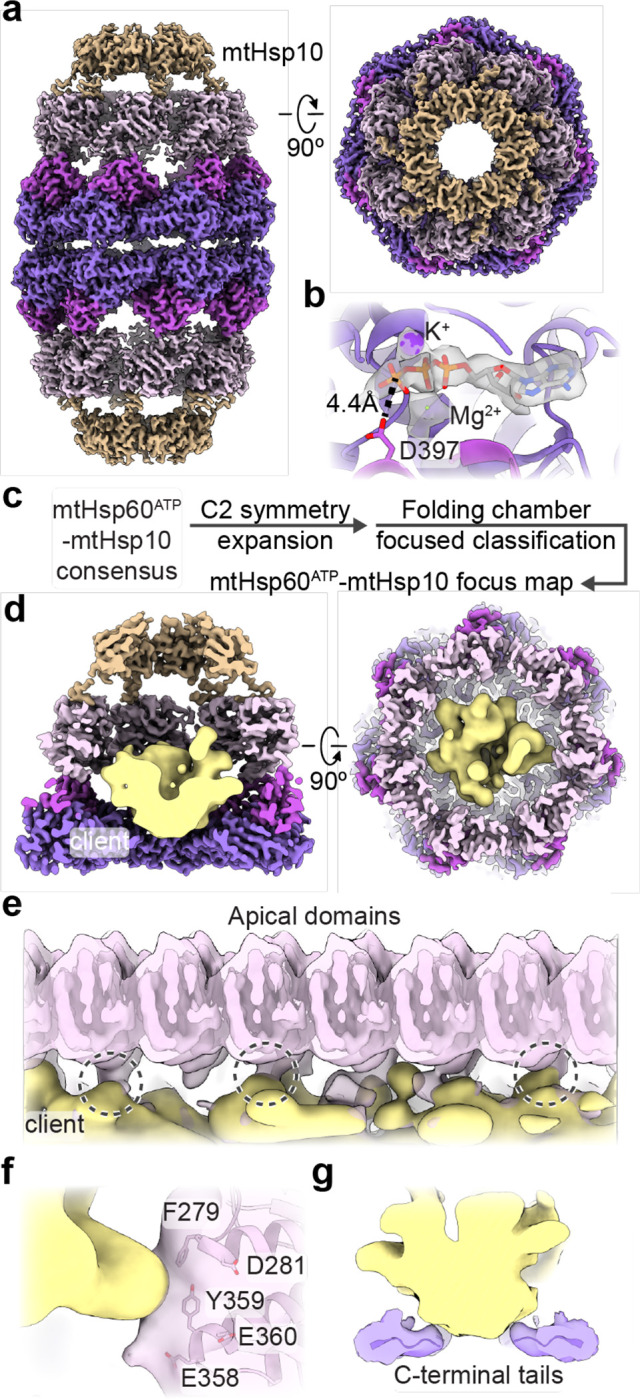
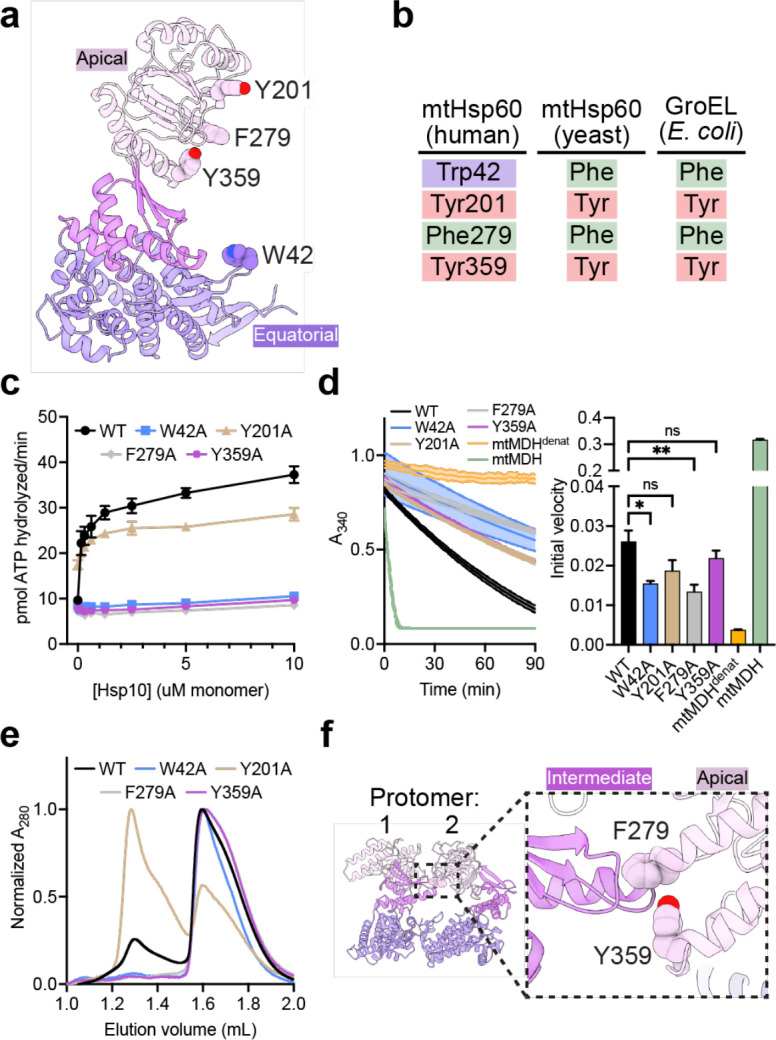
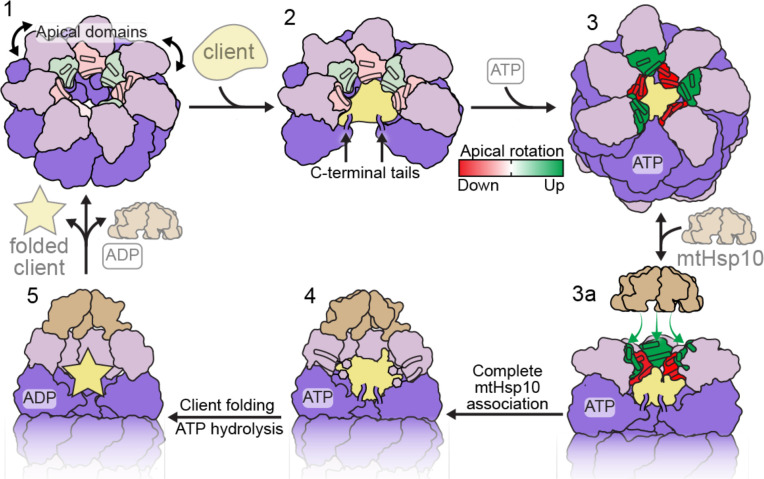
The mitochondrial chaperonin, mtHsp60, promotes the folding of newly imported and transiently misfolded proteins in the mitochondrial matrix, assisted by its co-chaperone mtHsp10. Despite its essential role in mitochondrial proteostasis, structural insights into how this chaperonin binds to clients and progresses through its ATP-dependent reaction cycle are not clear. Here, we determined cryo-electron microscopy (cryo-EM) structures of a hyperstable disease-associated mtHsp60 mutant, V72I, at three stages in this cycle. Unexpectedly, client density is identified in all states, revealing interactions with mtHsp60's apical domains and C-termini that coordinate client positioning in the folding chamber. We further identify a striking asymmetric arrangement of the apical domains in the ATP state, in which an alternating up/down configuration positions interaction surfaces for simultaneous recruitment of mtHsp10 and client retention. Client is then fully encapsulated in mtHsp60/mtHsp10, revealing prominent contacts at two discrete sites that potentially support maturation. These results identify a new role for the apical domains in coordinating client capture and progression through the cycle, and suggest a conserved mechanism of group I chaperonin function.
Conflict of interest statement
DECLARATION OF INTERESTS: The authors declare no competing interests.
Figures

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous