

Poly(A)-binding protein is an ataxin-2 chaperone that regulates biomolecular condensates
- PMID: 37295429
- PMCID: PMC10318123
- DOI: 10.1016/j.molcel.2023.05.025
Poly(A)-binding protein is an ataxin-2 chaperone that regulates biomolecular condensates
Abstract
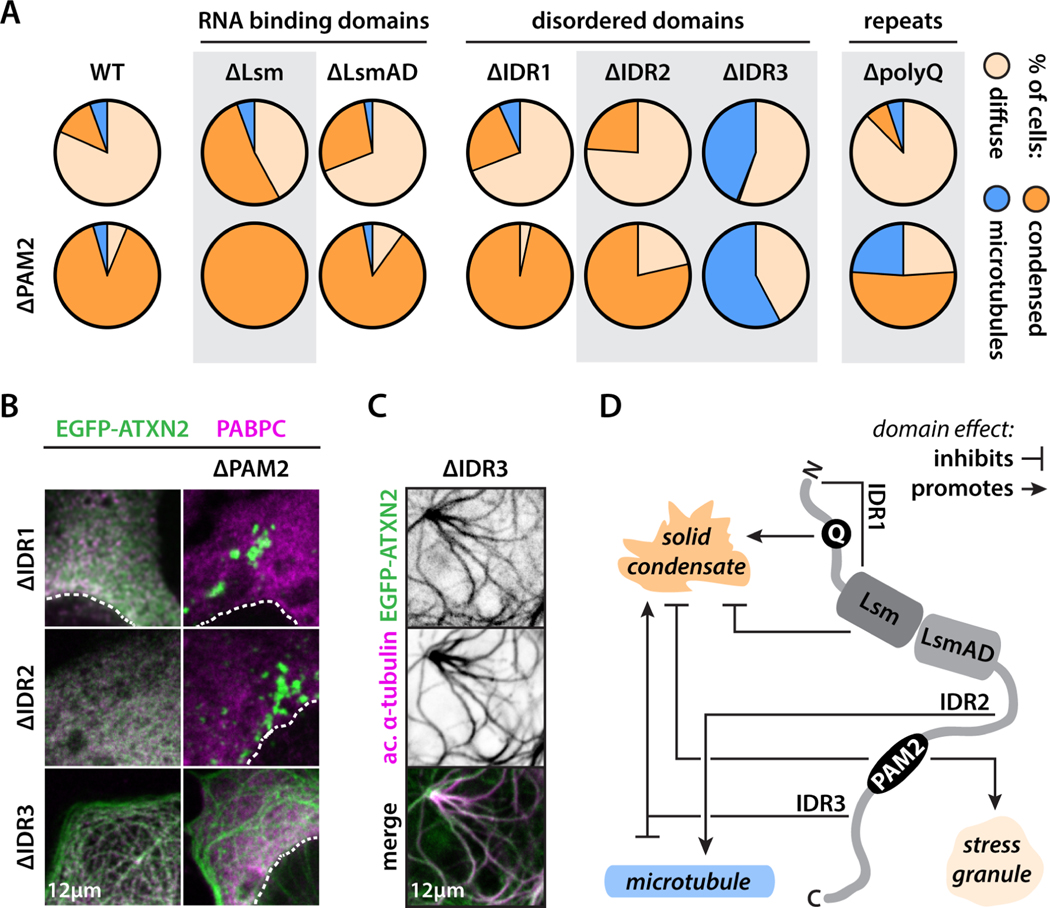
Biomolecular condensation underlies the biogenesis of an expanding array of membraneless assemblies, including stress granules (SGs), which form under a variety of cellular stresses. Advances have been made in understanding the molecular grammar of a few scaffold proteins that make up these phases, but how the partitioning of hundreds of SG proteins is regulated remains largely unresolved. While investigating the rules that govern the condensation of ataxin-2, an SG protein implicated in neurodegenerative disease, we unexpectedly identified a short 14 aa sequence that acts as a condensation switch and is conserved across the eukaryote lineage. We identify poly(A)-binding proteins as unconventional RNA-dependent chaperones that control this regulatory switch. Our results uncover a hierarchy of cis and trans interactions that fine-tune ataxin-2 condensation and reveal an unexpected molecular function for ancient poly(A)-binding proteins as regulators of biomolecular condensate proteins. These findings may inspire approaches to therapeutically target aberrant phases in disease.
Keywords: ATXN2; PABPC; amyotrophic lateral sclerosis; microtubule-binding protein; polyQ; protein aggregation; protein phase separation; short linear motif; spinocerebellar ataxia; stress granules.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.D.G. is a scientific founder of Maze Therapeutics. A.S.H. is a scientific consultant for Dewpoint Therapeutics and on the Scientific Advisory Board for Prose Foods.
Figures





Comment in
-
Stressing the role of a short linear motif in ataxin-2 condensation.Mol Cell. 2023 Jun 15;83(12):1961-1963. doi: 10.1016/j.molcel.2023.05.024. Mol Cell. 2023. PMID: 37327772
References
-
- McGurk L, Gomes E, Guo L, Mojsilovic-Petrovic J, Tran V, Kalb RG, Shorter J, and Bonini NM (2018). Poly(ADP-Ribose) Prevents Pathological Phase Separation of TDP-43 by Promoting Liquid Demixing and Stress Granule Localization. Mol Cell 71, 703–717 e709. 10.1016/j.molcel.2018.07.002. - DOI - PMC - PubMed
-
- Guillen-Boixet J, Kopach A, Holehouse AS, Wittmann S, Jahnel M, Schlussler R, Kim K, Trussina I, Wang J, Mateju D, et al. (2020). RNA-Induced Conformational Switching and Clustering of G3BP Drive Stress Granule Assembly by Condensation. Cell 181, 346–361 e317. 10.1016/j.cell.2020.03.049. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
