

Quorum sensing as a mechanism to harness the wisdom of the crowds
- PMID: 37296108
- PMCID: PMC10256802
- DOI: 10.1038/s41467-023-37950-7
Quorum sensing as a mechanism to harness the wisdom of the crowds
Abstract
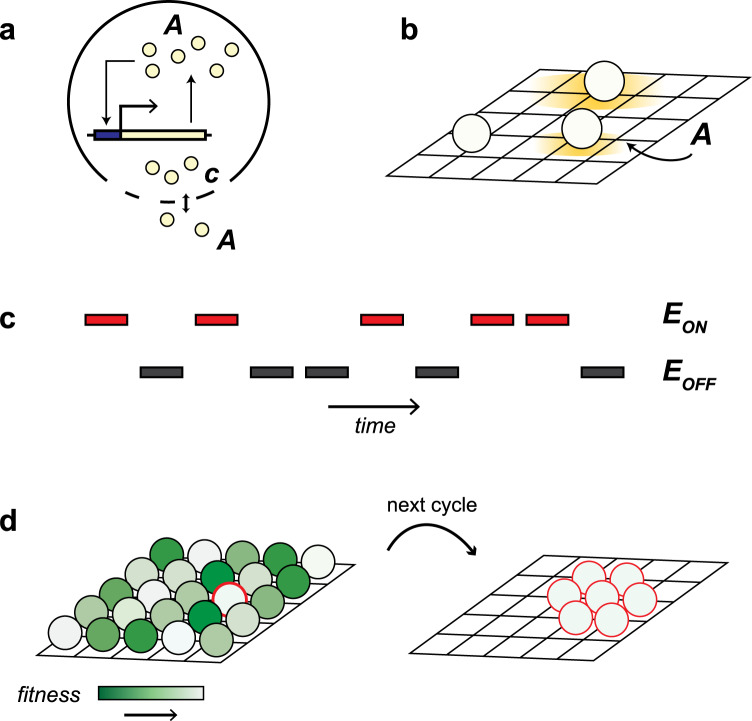
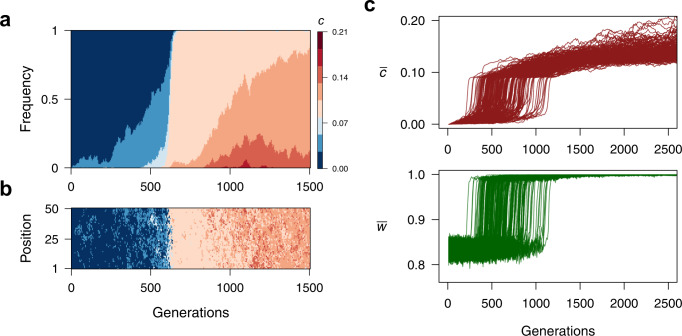
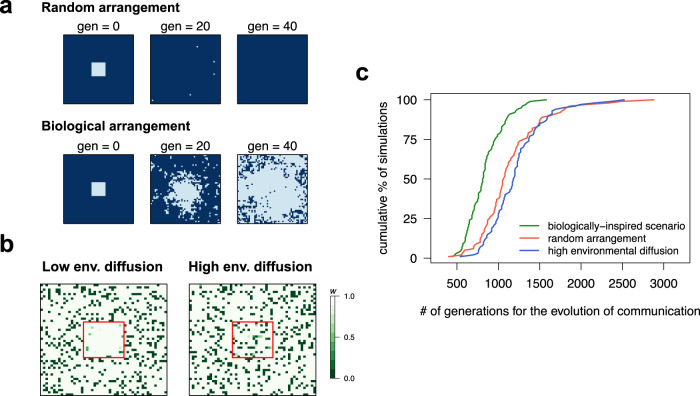
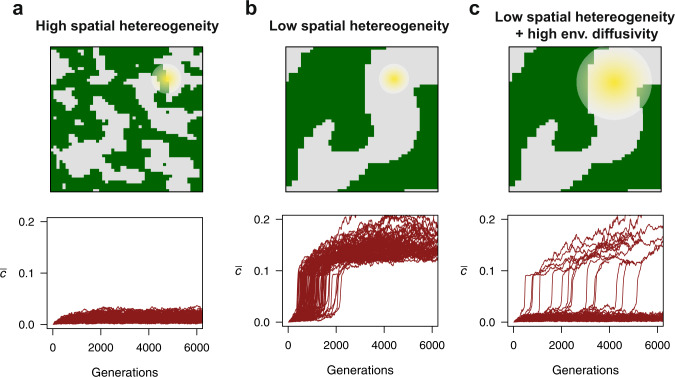
Bacteria release and sense small molecules called autoinducers in a process known as quorum sensing. The prevailing interpretation of quorum sensing is that by sensing autoinducer concentrations, bacteria estimate population density to regulate the expression of functions that are only beneficial when carried out by a sufficiently large number of cells. However, a major challenge to this interpretation is that the concentration of autoinducers strongly depends on the environment, often rendering autoinducer-based estimates of cell density unreliable. Here we propose an alternative interpretation of quorum sensing, where bacteria, by releasing and sensing autoinducers, harness social interactions to sense the environment as a collective. Using a computational model we show that this functionality can explain the evolution of quorum sensing and arises from individuals improving their estimation accuracy by pooling many imperfect estimates - analogous to the 'wisdom of the crowds' in decision theory. Importantly, our model reconciles the observed dependence of quorum sensing on both population density and the environment and explains why several quorum sensing systems regulate the production of private goods.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Saccharomyces cerevisiae Requires CFF1 To Produce 4-Hydroxy-5-Methylfuran-3(2H)-One, a Mimic of the Bacterial Quorum-Sensing Autoinducer AI-2.mBio. 2021 Mar 9;12(2):e03303-20. doi: 10.1128/mBio.03303-20. mBio. 2021. PMID: 33688008 Free PMC article.
-
Ecological feedback in quorum-sensing microbial populations can induce heterogeneous production of autoinducers.Elife. 2017 Jul 25;6:e25773. doi: 10.7554/eLife.25773. Elife. 2017. PMID: 28741470 Free PMC article.
-
The Vibrio cholerae Quorum-Sensing Protein VqmA Integrates Cell Density, Environmental, and Host-Derived Cues into the Control of Virulence.mBio. 2020 Jul 28;11(4):e01572-20. doi: 10.1128/mBio.01572-20. mBio. 2020. PMID: 32723922 Free PMC article.
-
Quorum Sensing Regulation as a Target for Antimicrobial Therapy.Mini Rev Med Chem. 2022;22(6):848-864. doi: 10.2174/1389557521666211202115259. Mini Rev Med Chem. 2022. PMID: 34856897 Review.
-
Quorum sensing in bacterial virulence.Microbiology (Reading). 2010 Aug;156(Pt 8):2271-2282. doi: 10.1099/mic.0.038794-0. Epub 2010 May 20. Microbiology (Reading). 2010. PMID: 20488878 Review.
Cited by
-
Emerging advances in biosensor technologies for quorum sensing signal molecules.Anal Bioanal Chem. 2025 Jan;417(1):33-50. doi: 10.1007/s00216-024-05659-1. Epub 2024 Nov 29. Anal Bioanal Chem. 2025. PMID: 39609273 Review.
-
Modeling bacterial interactions uncovers the importance of outliers in the coastal lignin-degrading consortium.Nat Commun. 2025 Jan 14;16(1):639. doi: 10.1038/s41467-025-56012-8. Nat Commun. 2025. PMID: 39809803 Free PMC article.
-
Physical communication pathways in bacteria: an extra layer to quorum sensing.Biophys Rev. 2025 Mar 4;17(2):667-685. doi: 10.1007/s12551-025-01290-1. eCollection 2025 Apr. Biophys Rev. 2025. PMID: 40376406 Free PMC article. Review.
-
Coumarins: Quorum Sensing and Biofilm Formation Inhibition.Molecules. 2024 Sep 24;29(19):4534. doi: 10.3390/molecules29194534. Molecules. 2024. PMID: 39407465 Free PMC article. Review.
-
DNA-programmed responsive microorganism assembly with controlled patterns and behaviors.Sci Adv. 2025 Jun 13;11(24):eads8651. doi: 10.1126/sciadv.ads8651. Epub 2025 Jun 13. Sci Adv. 2025. PMID: 40512851 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
