

In vivo PAR-CLIP (viP-CLIP) of liver TIAL1 unveils targets regulating cholesterol synthesis and secretion
- PMID: 37296170
- PMCID: PMC10256721
- DOI: 10.1038/s41467-023-39135-8
In vivo PAR-CLIP (viP-CLIP) of liver TIAL1 unveils targets regulating cholesterol synthesis and secretion
Abstract
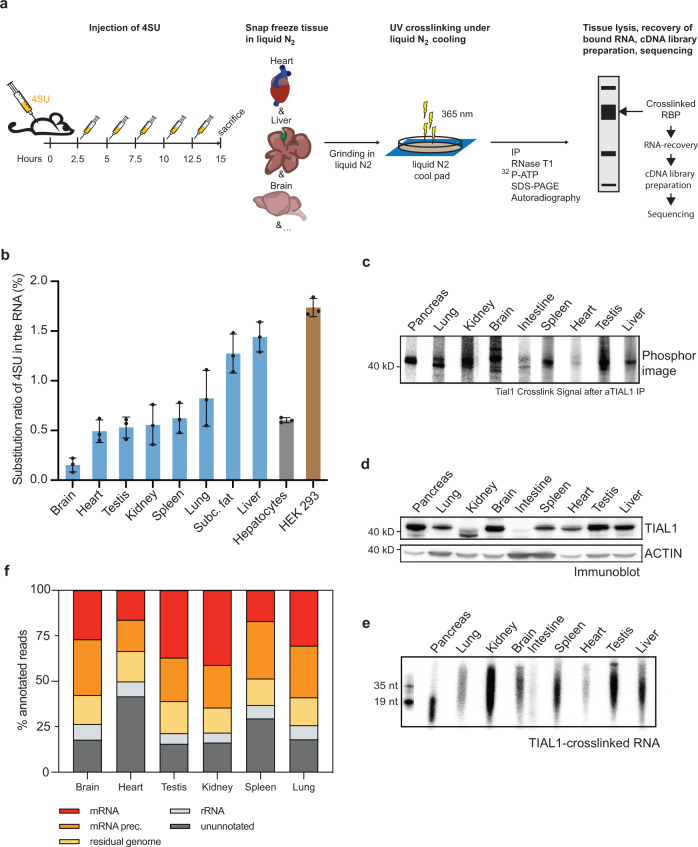
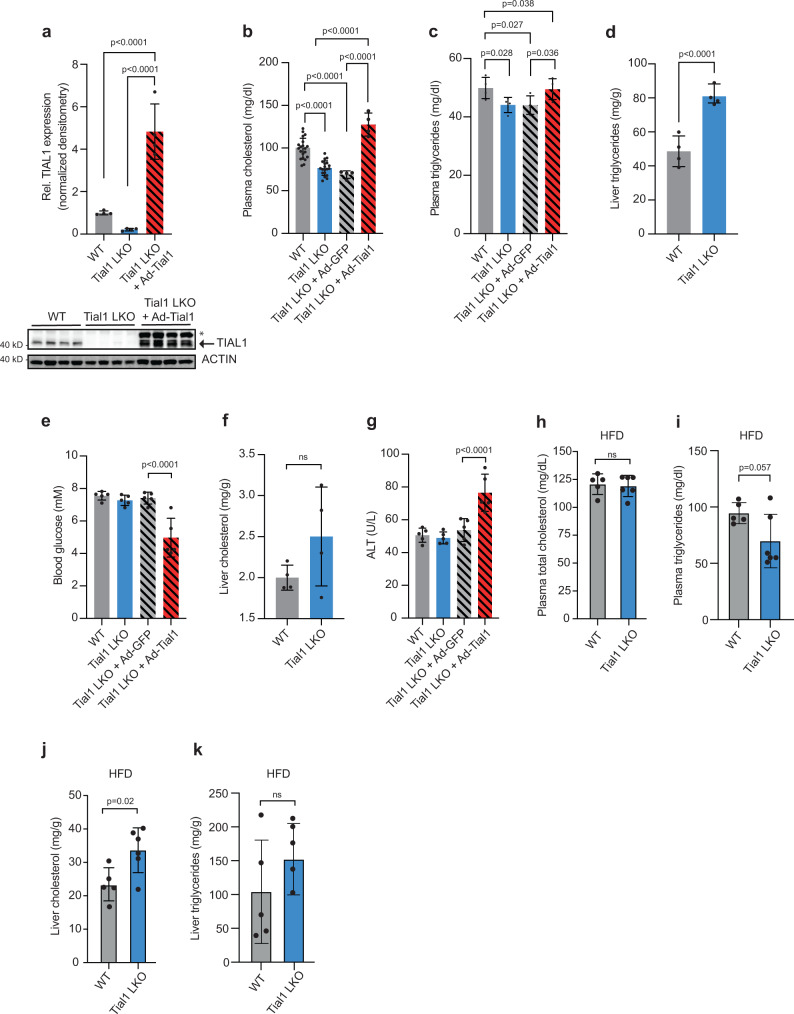
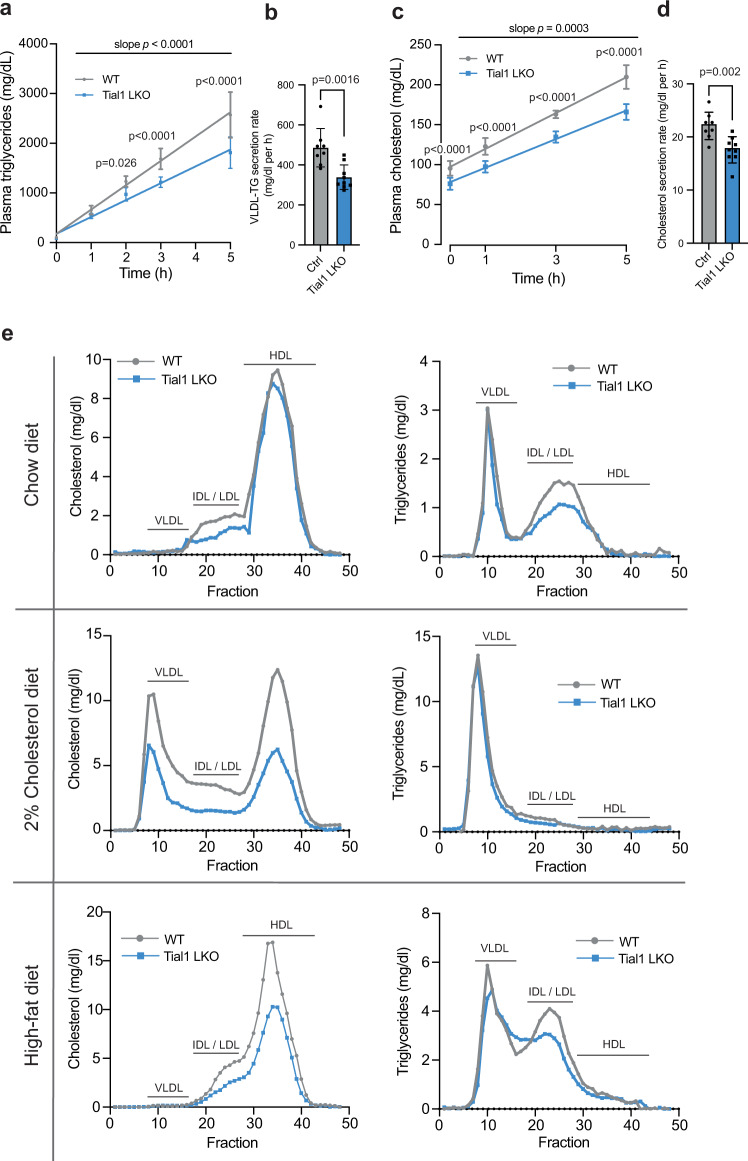
System-wide cross-linking and immunoprecipitation (CLIP) approaches have unveiled regulatory mechanisms of RNA-binding proteins (RBPs) mainly in cultured cells due to limitations in the cross-linking efficiency of tissues. Here, we describe viP-CLIP (in vivo PAR-CLIP), a method capable of identifying RBP targets in mammalian tissues, thereby facilitating the functional analysis of RBP-regulatory networks in vivo. We applied viP-CLIP to mouse livers and identified Insig2 and ApoB as prominent TIAL1 target transcripts, indicating an important role of TIAL1 in cholesterol synthesis and secretion. The functional relevance of these targets was confirmed by showing that TIAL1 influences their translation in hepatocytes. Mutant Tial1 mice exhibit altered cholesterol synthesis, APOB secretion and plasma cholesterol levels. Our results demonstrate that viP-CLIP can identify physiologically relevant RBP targets by finding a factor implicated in the negative feedback regulation of cholesterol biosynthesis.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
PAR-CliP--a method to identify transcriptome-wide the binding sites of RNA binding proteins.J Vis Exp. 2010 Jul 2;(41):2034. doi: 10.3791/2034. J Vis Exp. 2010. PMID: 20644507 Free PMC article.
-
Transcriptome-wide Identification of RNA-binding Protein Binding Sites Using Photoactivatable-Ribonucleoside-Enhanced Crosslinking Immunoprecipitation (PAR-CLIP).Curr Protoc Mol Biol. 2017 Apr 3;118:27.6.1-27.6.19. doi: 10.1002/cpmb.35. Curr Protoc Mol Biol. 2017. PMID: 28369676
-
Human protein-RNA interaction network is highly stable across mammals.BMC Genomics. 2019 Dec 30;20(Suppl 12):1004. doi: 10.1186/s12864-019-6330-9. BMC Genomics. 2019. PMID: 31888461 Free PMC article.
-
[Advances in technologies for large-scale enrichment and identification of ribonucleic acid-protein complexes].Se Pu. 2021 Feb;39(2):105-111. doi: 10.3724/SP.J.1123.2020.07019. Se Pu. 2021. PMID: 34227341 Free PMC article. Review. Chinese.
-
Identification of RNA-protein interaction networks using PAR-CLIP.Wiley Interdiscip Rev RNA. 2012 Mar-Apr;3(2):159-77. doi: 10.1002/wrna.1103. Epub 2011 Dec 27. Wiley Interdiscip Rev RNA. 2012. PMID: 22213601 Free PMC article. Review.
Cited by
-
Alternative splicing and related RNA binding proteins in human health and disease.Signal Transduct Target Ther. 2024 Feb 2;9(1):26. doi: 10.1038/s41392-024-01734-2. Signal Transduct Target Ther. 2024. PMID: 38302461 Free PMC article. Review.
-
PAR-dCLIP: Enabling detection of RNA binding protein target transcripts bound at 5' termini through the incorporation of a decapping step.Methods Enzymol. 2024;705:159-222. doi: 10.1016/bs.mie.2024.08.003. Epub 2024 Sep 7. Methods Enzymol. 2024. PMID: 39389663 Free PMC article.
-
Molecular Regulation and Therapeutic Targeting of VLDL Production in Cardiometabolic Disease.Cell Mol Gastroenterol Hepatol. 2025;19(1):101409. doi: 10.1016/j.jcmgh.2024.101409. Epub 2024 Oct 12. Cell Mol Gastroenterol Hepatol. 2025. PMID: 39406347 Free PMC article. Review.
-
VLDL Biogenesis and Secretion: It Takes a Village.Circ Res. 2024 Jan 19;134(2):226-244. doi: 10.1161/CIRCRESAHA.123.323284. Epub 2024 Jan 18. Circ Res. 2024. PMID: 38236950 Free PMC article. Review.
References
-
- Hafner M, et al. CLIP and complementary methods. Nat. Rev. Methods Prim. 2021;1:1–23.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
