Whole genome analysis for 163 gRNAs in Cas9-edited mice reveals minimal off-target activity
- PMID: 37301944
- PMCID: PMC10257658
- DOI: 10.1038/s42003-023-04974-0
Whole genome analysis for 163 gRNAs in Cas9-edited mice reveals minimal off-target activity
Abstract
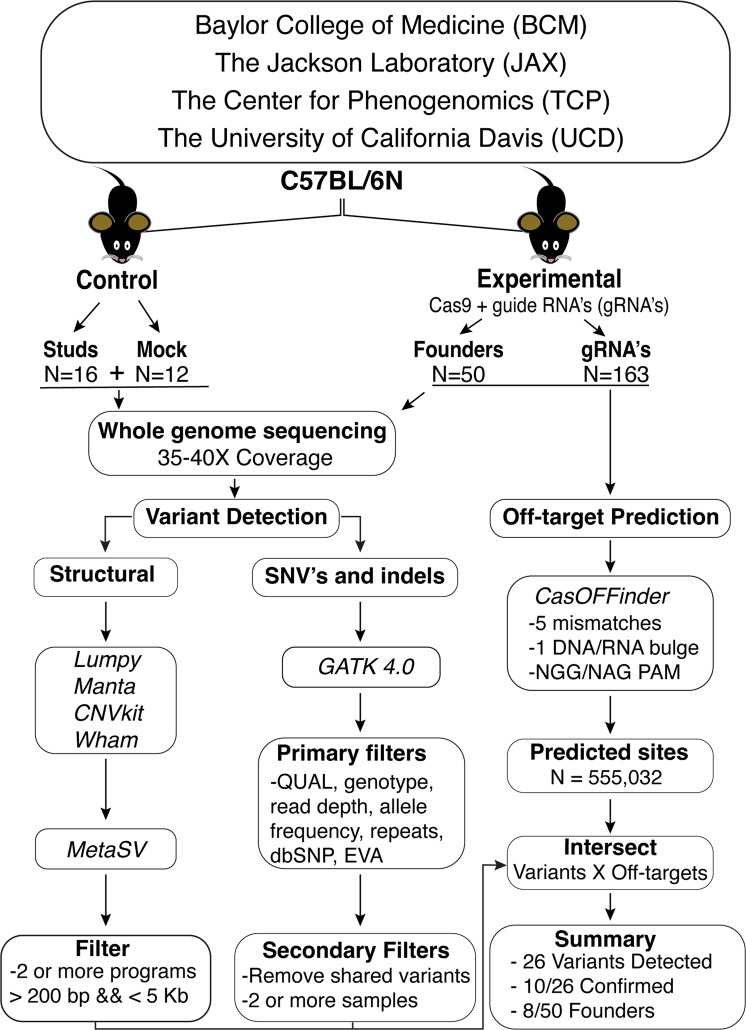
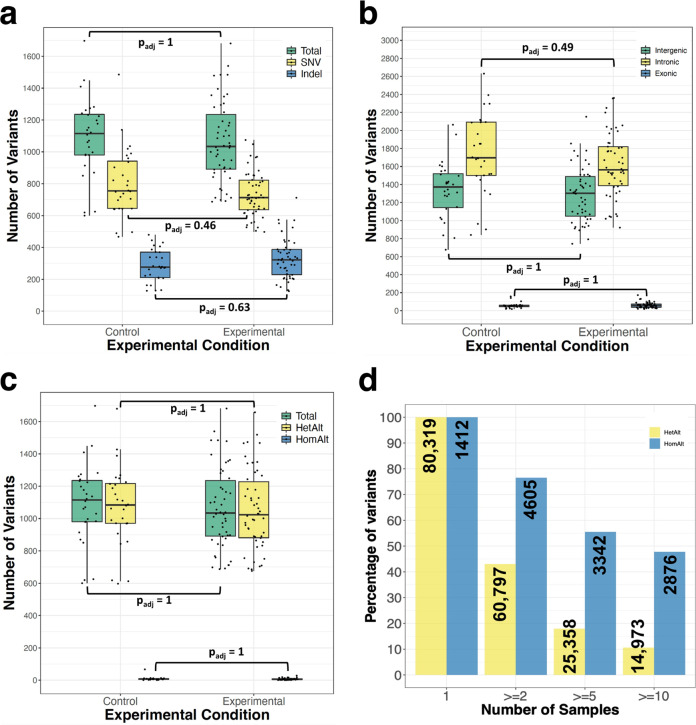
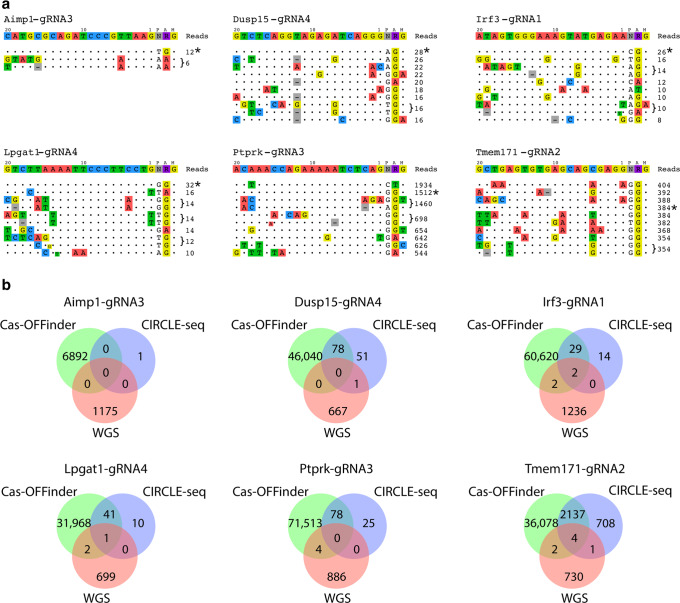
Genome editing with CRISPR-associated (Cas) proteins holds exceptional promise for "correcting" variants causing genetic disease. To realize this promise, off-target genomic changes cannot occur during the editing process. Here, we use whole genome sequencing to compare the genomes of 50 Cas9-edited founder mice to 28 untreated control mice to assess the occurrence of S. pyogenes Cas9-induced off-target mutagenesis. Computational analysis of whole-genome sequencing data detects 26 unique sequence variants at 23 predicted off-target sites for 18/163 guides used. While computationally detected variants are identified in 30% (15/50) of Cas9 gene-edited founder animals, only 38% (10/26) of the variants in 8/15 founders validate by Sanger sequencing. In vitro assays for Cas9 off-target activity identify only two unpredicted off-target sites present in genome sequencing data. In total, only 4.9% (8/163) of guides tested have detectable off-target activity, a rate of 0.2 Cas9 off-target mutations per founder analyzed. In comparison, we observe ~1,100 unique variants in each mouse regardless of genome exposure to Cas9 indicating off-target variants comprise a small fraction of genetic heterogeneity in Cas9-edited mice. These findings will inform future design and use of Cas9-edited animal models as well as provide context for evaluating off-target potential in genetically diverse patient populations.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures