

Chronic alcohol consumption shifts learning strategies and synaptic plasticity from hippocampus to striatum-dependent pathways
- PMID: 37304443
- PMCID: PMC10250670
- DOI: 10.3389/fpsyt.2023.1129030
Chronic alcohol consumption shifts learning strategies and synaptic plasticity from hippocampus to striatum-dependent pathways
Abstract
Introduction: The hippocampus and striatum have dissociable roles in memory and are necessary for spatial and procedural/cued learning, respectively. Emotionally charged, stressful events promote the use of striatal- over hippocampus-dependent learning through the activation of the amygdala. An emerging hypothesis suggests that chronic consumption of addictive drugs similarly disrupt spatial/declarative memory while facilitating striatum-dependent associative learning. This cognitive imbalance could contribute to maintain addictive behaviors and increase the risk of relapse.
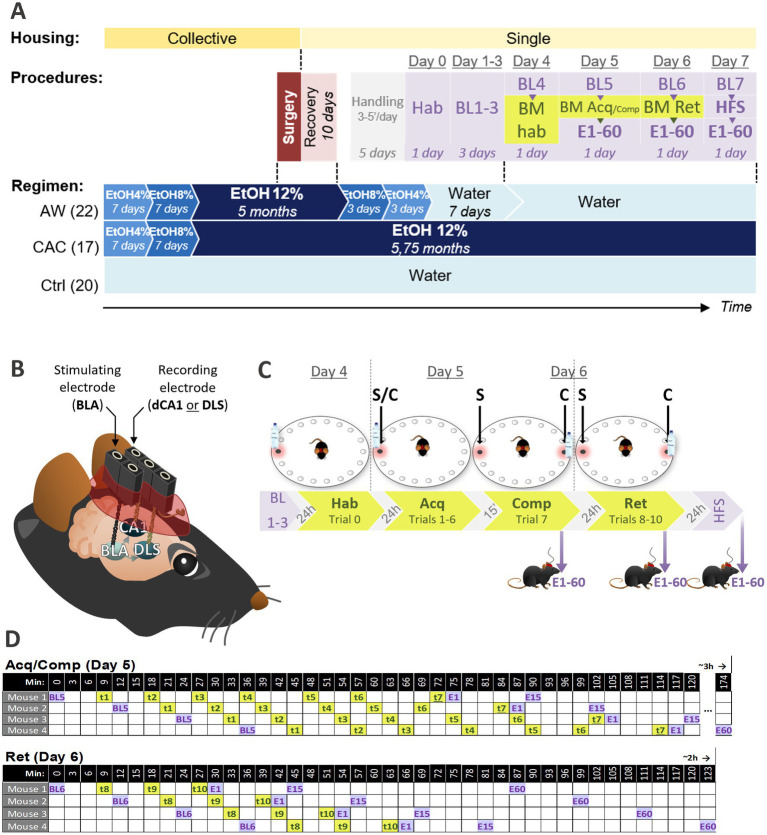
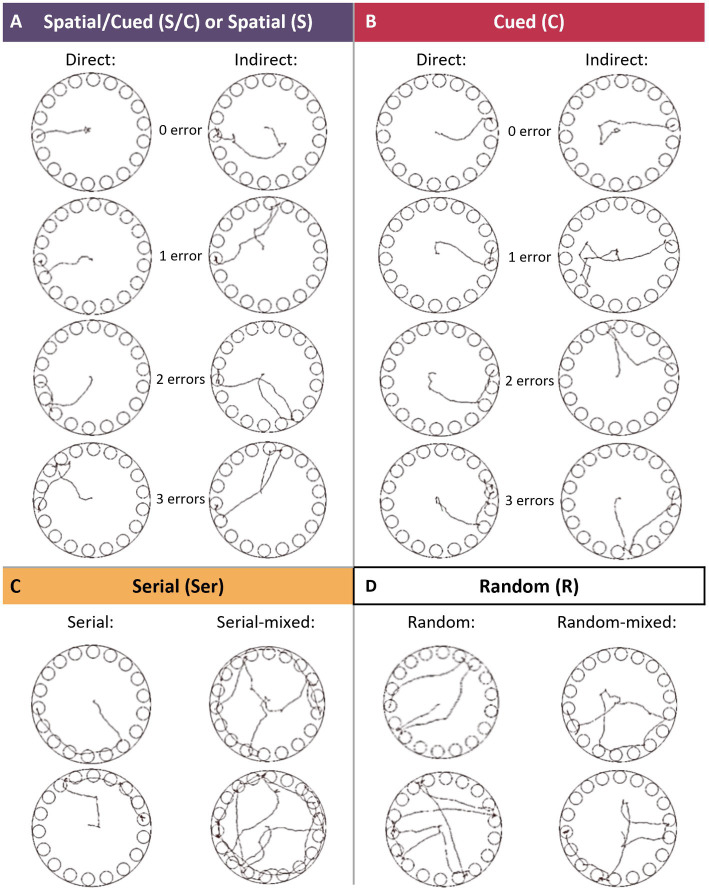
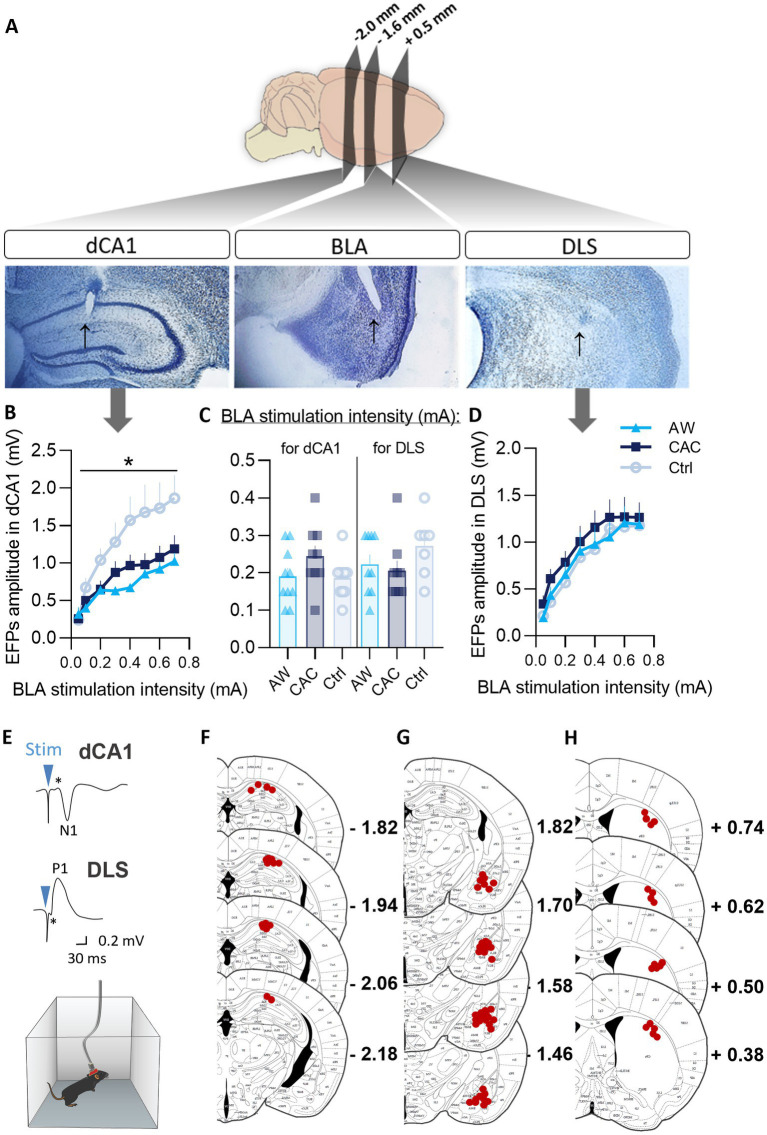
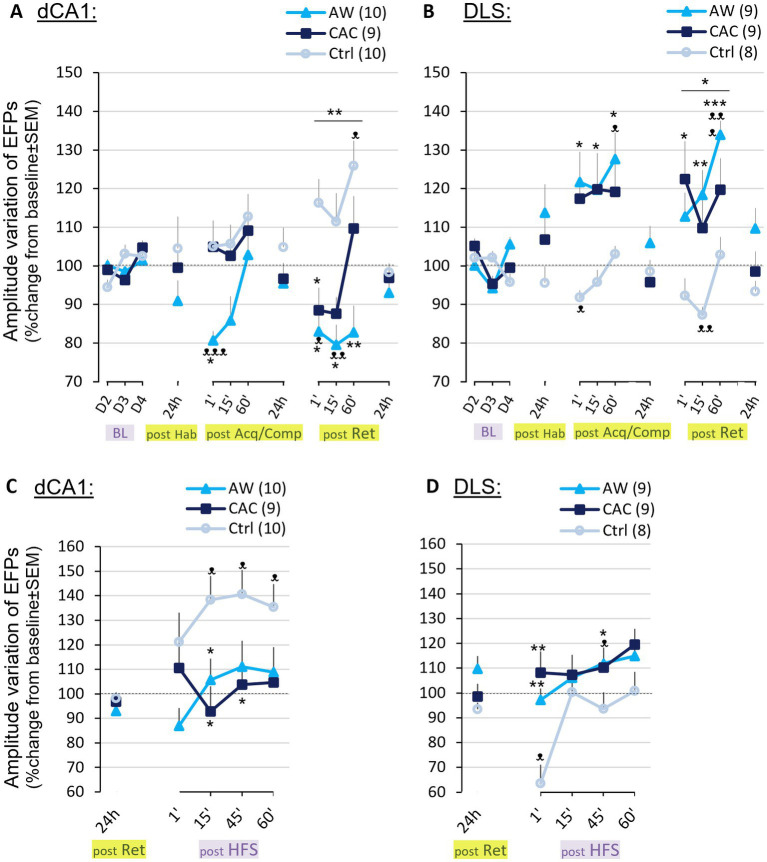
Methods: We first examined, in C57BL/6 J male mice, whether chronic alcohol consumption (CAC) and alcohol withdrawal (AW) might modulate the respective use of spatial vs. single cue-based learning strategies, using a competition protocol in the Barnes maze task. We then performed in vivo electrophysiological studies in freely moving mice to assess learning-induced synaptic plasticity in both the basolateral amygdala (BLA) to dorsal hippocampus (dCA1) and BLA to dorsolateral striatum (DLS) pathways.
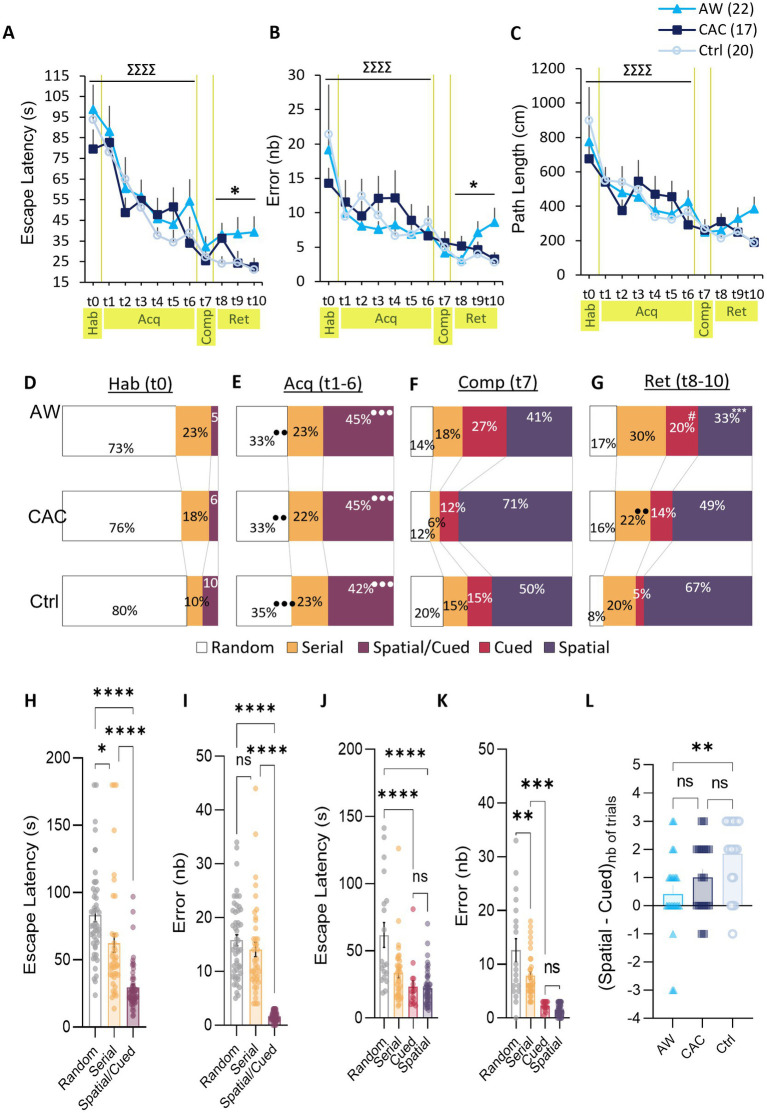
Results: We found that both CAC and early AW promote the use of cue-dependent learning strategies, and potentiate plasticity in the BLA → DLS pathway while reducing the use of spatial memory and depressing BLA → dCA1 neurotransmission.
Discussion: These results support the view that CAC disrupt normal hippocampo-striatal interactions, and suggest that targeting this cognitive imbalance through spatial/declarative task training could be of great help to maintain protracted abstinence in alcoholic patients.
Keywords: addiction; alcohol; amygdala; dorsal striatum; hippocampus (CA1); learning strategies; memory systems; synaptic plasticity.
Copyright © 2023 Tochon, Vouimba, Corio, Henkous, Béracochéa, Guillou and David.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
The medial entorhinal cortex mediates basolateral amygdala effects on spatial memory and downstream activity-regulated cytoskeletal-associated protein expression.Neuropsychopharmacology. 2021 May;46(6):1172-1182. doi: 10.1038/s41386-020-00875-6. Epub 2020 Oct 2. Neuropsychopharmacology. 2021. PMID: 33007779 Free PMC article.
-
The amygdala and emotional modulation of competition between cognitive and habit memory.Behav Brain Res. 2008 Nov 3;193(1):126-31. doi: 10.1016/j.bbr.2008.05.002. Epub 2008 May 8. Behav Brain Res. 2008. PMID: 18565602
-
Basolateral Amygdala Inputs to the Medial Entorhinal Cortex Selectively Modulate the Consolidation of Spatial and Contextual Learning.J Neurosci. 2018 Mar 14;38(11):2698-2712. doi: 10.1523/JNEUROSCI.2848-17.2018. Epub 2018 Feb 5. J Neurosci. 2018. PMID: 29431646 Free PMC article.
-
Amygdala-hippocampal interactions in synaptic plasticity and memory formation.Neurobiol Learn Mem. 2021 Oct;184:107490. doi: 10.1016/j.nlm.2021.107490. Epub 2021 Jul 21. Neurobiol Learn Mem. 2021. PMID: 34302951 Free PMC article. Review.
-
Memory Systems and the Addicted Brain.Front Psychiatry. 2016 Feb 25;7:24. doi: 10.3389/fpsyt.2016.00024. eCollection 2016. Front Psychiatry. 2016. PMID: 26941660 Free PMC article. Review.
Cited by
-
Impact and Interrelationships of Striatal Proteins, EPHB2, OPRM1, and PER2 on Mild Cognitive Impairment.Mol Neurobiol. 2025 Feb;62(2):1478-1492. doi: 10.1007/s12035-024-04334-x. Epub 2024 Jul 13. Mol Neurobiol. 2025. PMID: 39002057 Free PMC article.
References
-
- Organization WH . Global status report on alcohol and health (2018). World Health Organization. 2019.
LinkOut - more resources
Full Text Sources
Miscellaneous