

A nonclassically secreted effector of Magnaporthe oryzae targets host nuclei and plays important roles in fungal growth and plant infection
- PMID: 37306516
- PMCID: PMC10423324
- DOI: 10.1111/mpp.13356
A nonclassically secreted effector of Magnaporthe oryzae targets host nuclei and plays important roles in fungal growth and plant infection
Abstract
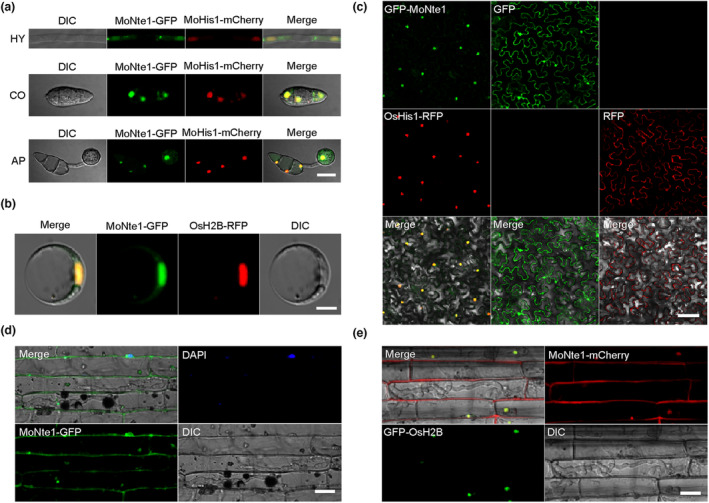
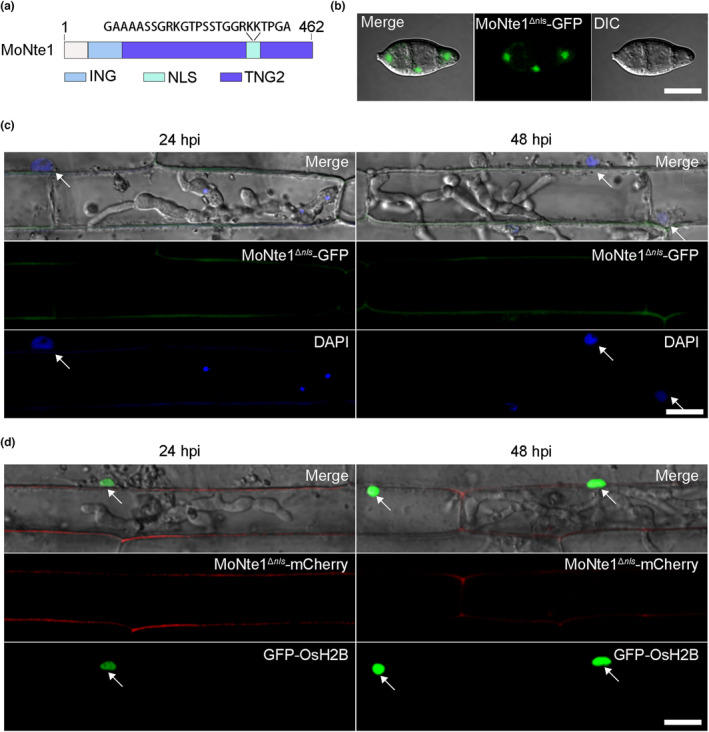
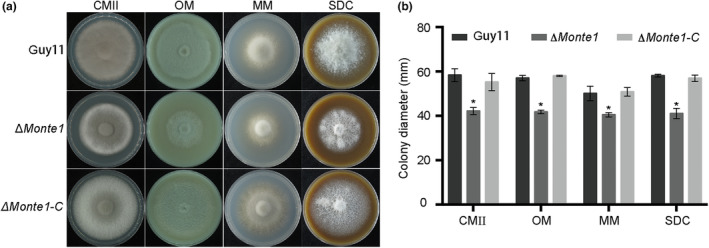
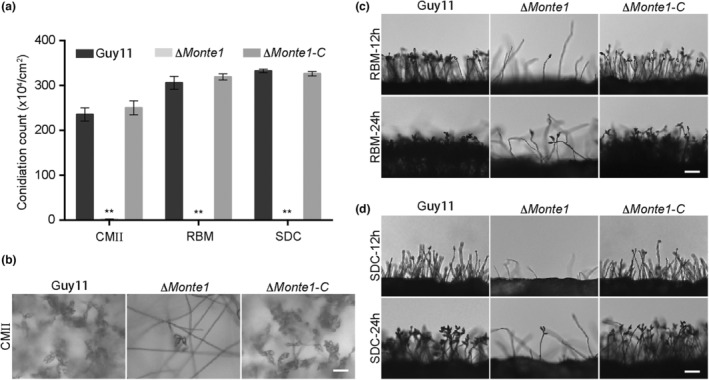
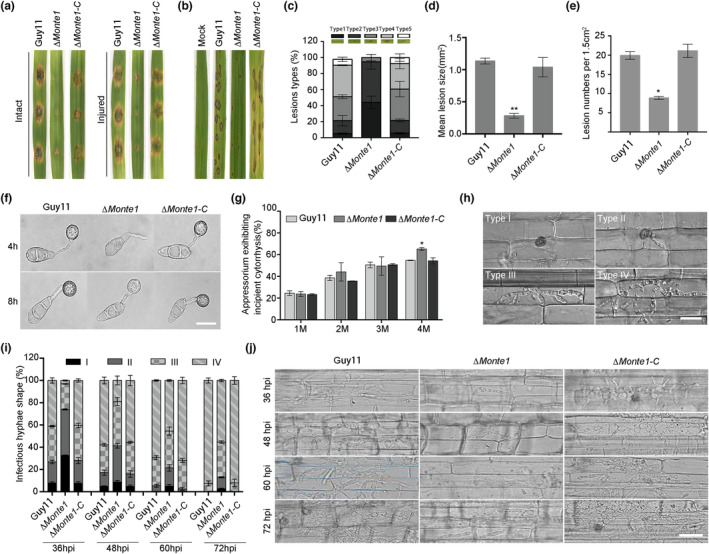
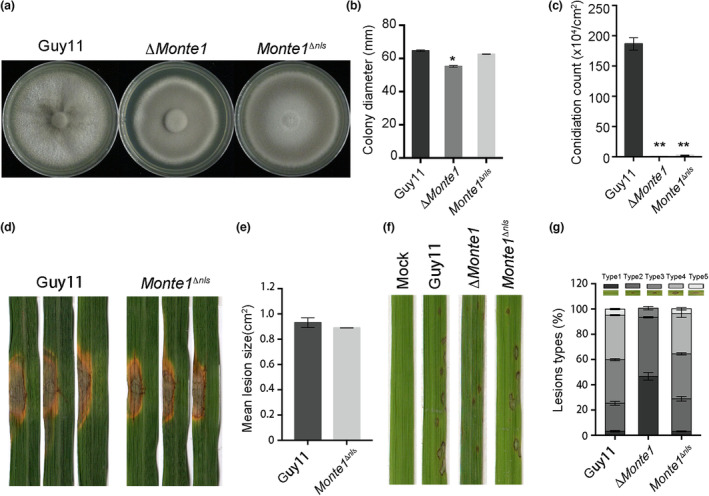
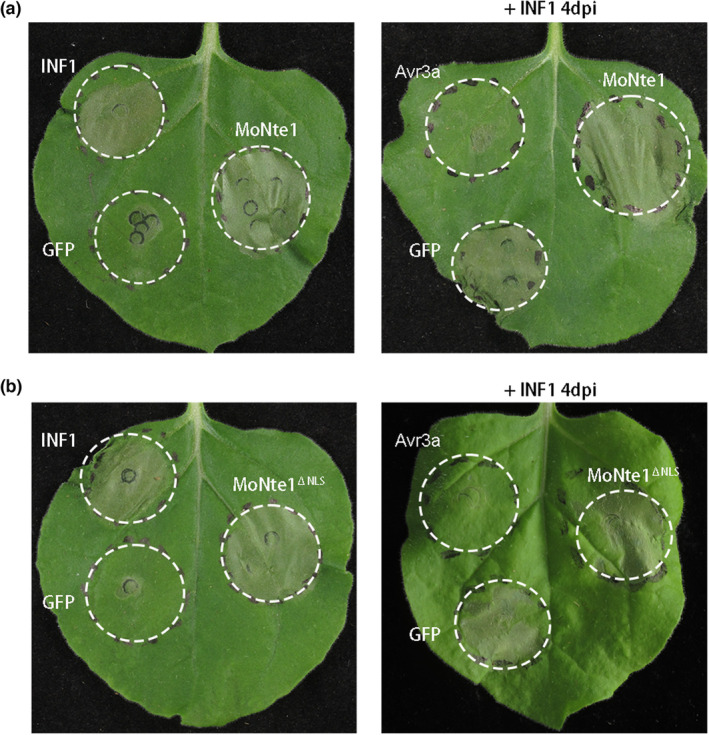
Rice blast caused by Magnaporthe oryzae is one of the most destructive diseases and poses a growing threat to food security worldwide. Like many other filamentous pathogens, rice blast fungus releases multiple types of effector proteins to facilitate fungal infection and modulate host defence responses. However, most of the characterized effectors contain an N-terminal signal peptide. Here, we report the results of the functional characterization of a nonclassically secreted nuclear targeting effector in M. oryzae (MoNte1). MoNte1 has no signal peptide, but can be secreted and translocated into plant nuclei driven by a nuclear targeting peptide. It could also induce hypersensitive cell death when transiently expressed in Nicotiana benthamiana. Deletion of the MoNTE1 gene caused a significant reduction of fungal growth and conidiogenesis, partially impaired appressorium formation and host colonization, and also dramatically attenuated the pathogenicity. Taken together, these findings reveal a novel effector secretion pathway and deepen our understanding of rice-M. oryzae interactions.
Keywords: Magnaporthe oryzae; fungal growth; hypersensitive cell death; nuclear targeting; pathogenicity; rice-Magnaporthe oryzae interactions; signal peptide.
© 2023 The Authors. Molecular Plant Pathology published by British Society for Plant Pathology and John Wiley & Sons Ltd.
Figures
Similar articles
-
Effector-triggered susceptibility by the rice blast fungus Magnaporthe oryzae.New Phytol. 2024 Feb;241(3):1007-1020. doi: 10.1111/nph.19446. Epub 2023 Dec 10. New Phytol. 2024. PMID: 38073141 Review.
-
The nuclear effector MoHTR3 of Magnaporthe oryzae modulates host defence signalling in the biotrophic stage of rice infection.Mol Plant Pathol. 2023 Jun;24(6):602-615. doi: 10.1111/mpp.13326. Epub 2023 Mar 28. Mol Plant Pathol. 2023. PMID: 36977203 Free PMC article.
-
A Histone Deacetylase, Magnaporthe oryzae RPD3, Regulates Reproduction and Pathogenic Development in the Rice Blast Fungus.mBio. 2021 Dec 21;12(6):e0260021. doi: 10.1128/mBio.02600-21. Epub 2021 Nov 16. mBio. 2021. PMID: 34781734 Free PMC article.
-
Large-scale gene disruption in Magnaporthe oryzae identifies MC69, a secreted protein required for infection by monocot and dicot fungal pathogens.PLoS Pathog. 2012;8(5):e1002711. doi: 10.1371/journal.ppat.1002711. Epub 2012 May 10. PLoS Pathog. 2012. PMID: 22589729 Free PMC article.
-
Investigating the cell and developmental biology of plant infection by the rice blast fungus Magnaporthe oryzae.Fungal Genet Biol. 2021 Sep;154:103562. doi: 10.1016/j.fgb.2021.103562. Epub 2021 Apr 18. Fungal Genet Biol. 2021. PMID: 33882359 Review.
Cited by
-
A Novel Effector FoUpe9 Enhances the Virulence of Fusarium oxysporum f. sp. cubense Tropical Race 4 by Inhibiting Plant Immunity.J Fungi (Basel). 2025 Apr 13;11(4):308. doi: 10.3390/jof11040308. J Fungi (Basel). 2025. PMID: 40278128 Free PMC article.
-
Plant Growth Promotion and Stress Tolerance Enhancement through Inoculation with Bacillus proteolyticus OSUB18.Biology (Basel). 2023 Dec 6;12(12):1495. doi: 10.3390/biology12121495. Biology (Basel). 2023. PMID: 38132321 Free PMC article.
-
The Xenopyricularia zizaniicola exhibits a genome architecture distinct to the two-speed genome.Microbiol Spectr. 2025 Jul;13(7):e0036225. doi: 10.1128/spectrum.00362-25. Epub 2025 Jun 9. Microbiol Spectr. 2025. PMID: 40488379 Free PMC article.
-
Wheat Leaf Rust Effector Pt48115 Localized in the Chloroplasts and Suppressed Wheat Immunity.J Fungi (Basel). 2025 Jan 20;11(1):80. doi: 10.3390/jof11010080. J Fungi (Basel). 2025. PMID: 39852499 Free PMC article.
-
The Essentials of PgPG1, a Polygalacturonase-Encoding Gene for the Invasion of Pyrenophora graminea to Hordeum vulgare.Int J Mol Sci. 2025 Mar 7;26(6):2401. doi: 10.3390/ijms26062401. Int J Mol Sci. 2025. PMID: 40141049 Free PMC article.
References
-
- Batool, W. , Shabbir, A. , Lin, L. , Chen, X. , An, Q. , He, X. et al. (2021) Translation initiation factor eIF4E positively modulates conidiogenesis, appressorium formation, host invasion and stress homeostasis in the filamentous fungi Magnaporthe oryzae . Frontiers in Plant Science, 12, 646343. - PMC - PubMed
-
- Boradia, V.M. , Malhotra, H. , Thakkar, J.S. , Tillu, V.A. , Vuppala, B. , Patil, P. et al. (2014) Mycobacterium tuberculosis acquires iron by cell‐surface sequestration and internalization of human holo‐transferrin. Nature Communications, 5, 54730. - PubMed
-
- Branco, P. , Francisco, D. , Chambon, C. , Hébraud, M. , Arneborg, N. , Almeida, M.G. et al. (2014) Identification of novel GAPDH‐derived antimicrobial peptides secreted by Saccharomyces cerevisiae and involved in wine microbial interactions. Applied Microbiology Biotechnology, 98, 843–853. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Research Materials
