

Dynamic chromatin architecture of the porcine adipose tissues with weight gain and loss
- PMID: 37308492
- PMCID: PMC10258790
- DOI: 10.1038/s41467-023-39191-0
Dynamic chromatin architecture of the porcine adipose tissues with weight gain and loss
Abstract
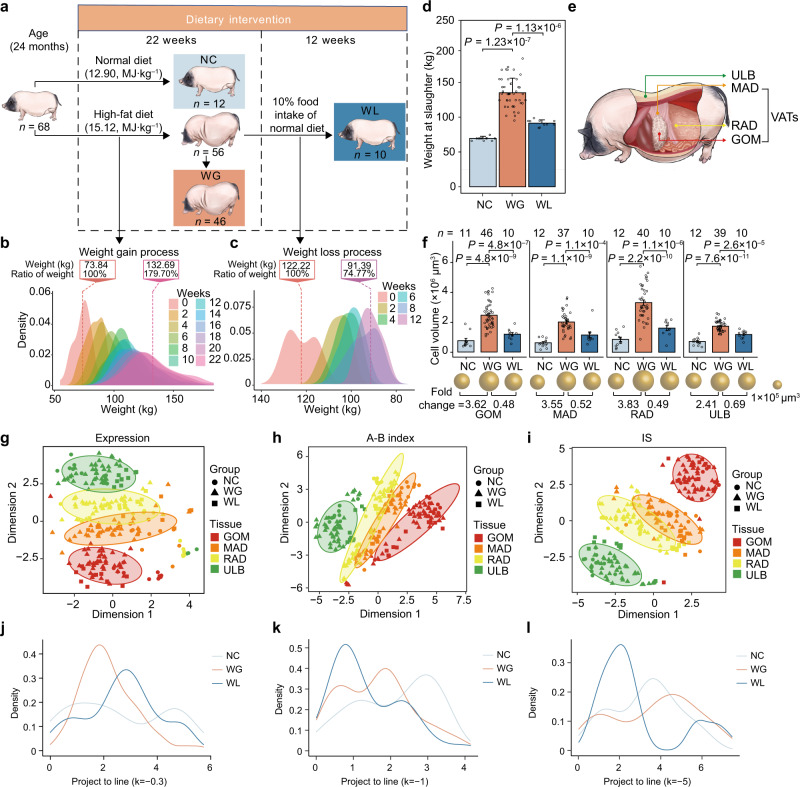
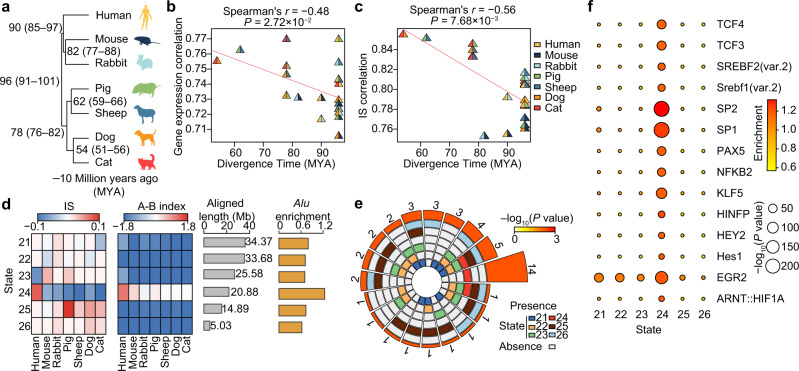
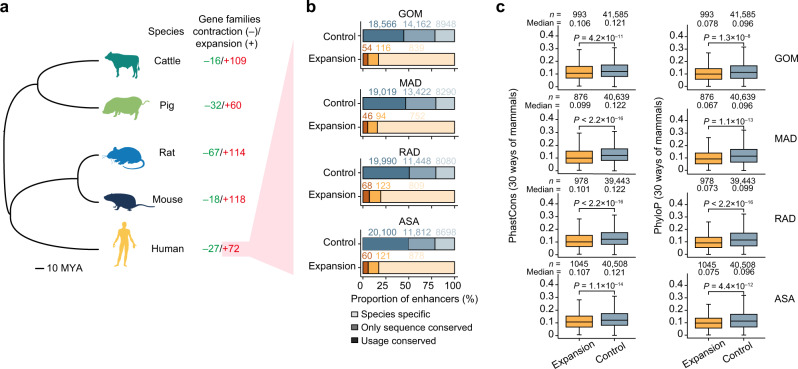
Using an adult female miniature pig model with diet-induced weight gain/weight loss, we investigated the regulatory mechanisms of three-dimensional (3D) genome architecture in adipose tissues (ATs) associated with obesity. We generated 249 high-resolution in situ Hi-C chromatin contact maps of subcutaneous AT and three visceral ATs, analyzing transcriptomic and chromatin architectural changes under different nutritional treatments. We find that chromatin architecture remodeling underpins transcriptomic divergence in ATs, potentially linked to metabolic risks in obesity development. Analysis of chromatin architecture among subcutaneous ATs of different mammals suggests the presence of transcriptional regulatory divergence that could explain phenotypic, physiological, and functional differences in ATs. Regulatory element conservation analysis in pigs and humans reveals similarities in the regulatory circuitry of genes responsible for the obesity phenotype and identified non-conserved elements in species-specific gene sets that underpin AT specialization. This work provides a data-rich tool for discovering obesity-related regulatory elements in humans and pigs.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- OECD/World Health Organization. Overweight and obesity. In Health at a Glance: Asia/Pacific 2020: Measuring Progress Towards Universal Health Coverage (OECD Publishing, 2020). https://www.oecd-ilibrary.org/social-issues-migration-health/health-at-a....
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
