

Transient water wires mediate selective proton transport in designed channel proteins
- PMID: 37308712
- PMCID: PMC10475958
- DOI: 10.1038/s41557-023-01210-4
Transient water wires mediate selective proton transport in designed channel proteins
Abstract
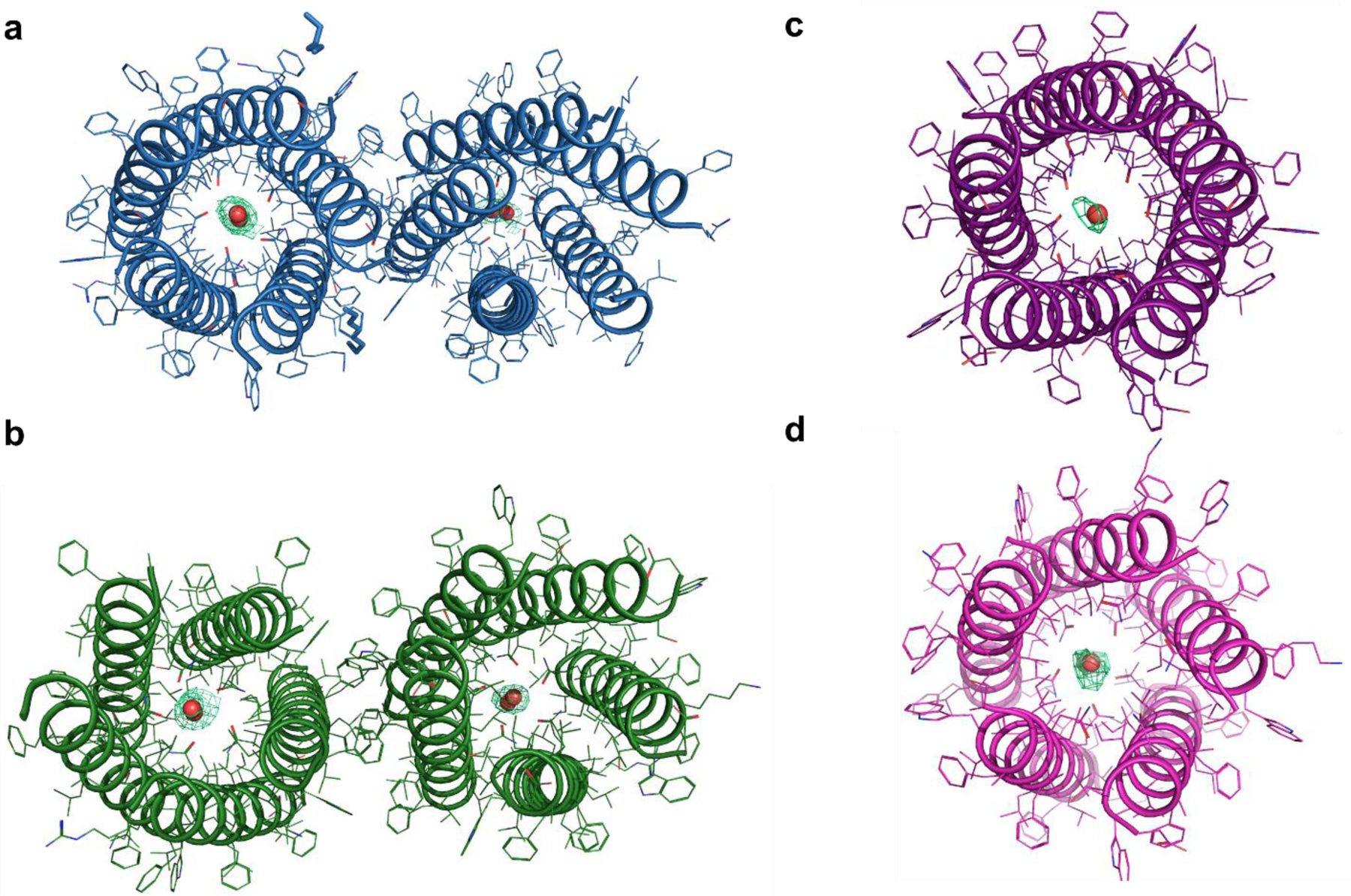
Selective proton transport through proteins is essential for forming and using proton gradients in cells. Protons are conducted along hydrogen-bonded 'wires' of water molecules and polar side chains, which, somewhat surprisingly, are often interrupted by dry apolar stretches in the conduction pathways, inferred from static protein structures. Here we hypothesize that protons are conducted through such dry spots by forming transient water wires, often highly correlated with the presence of the excess protons in the water wire. To test this hypothesis, we performed molecular dynamics simulations to design transmembrane channels with stable water pockets interspersed by apolar segments capable of forming flickering water wires. The minimalist designed channels conduct protons at rates similar to viral proton channels, and they are at least 106-fold more selective for H+ over Na+. These studies inform the mechanisms of biological proton conduction and the principles for engineering proton-conductive materials.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Figures













Similar articles
-
Molecular mechanism of H+ conduction in the single-file water chain of the gramicidin channel.Biophys J. 2002 May;82(5):2304-16. doi: 10.1016/S0006-3495(02)75576-8. Biophys J. 2002. PMID: 11964221 Free PMC article.
-
Hydrated Excess Protons Can Create Their Own Water Wires.J Phys Chem B. 2015 Jul 23;119(29):9212-8. doi: 10.1021/jp5095118. Epub 2014 Nov 12. J Phys Chem B. 2015. PMID: 25369445 Free PMC article.
-
Free energy profiles for H+ conduction along hydrogen-bonded chains of water molecules.Biophys J. 1998 Jul;75(1):33-40. doi: 10.1016/S0006-3495(98)77492-2. Biophys J. 1998. PMID: 9649365 Free PMC article.
-
The transfer of protons in water wires inside proteins.Front Biosci. 2003 Sep 1;8:s1118-39. doi: 10.2741/1192. Front Biosci. 2003. PMID: 12957840 Review.
-
Proton permeation of lipid bilayers.J Bioenerg Biomembr. 1987 Oct;19(5):457-79. doi: 10.1007/BF00770030. J Bioenerg Biomembr. 1987. PMID: 2447068 Review.
Cited by
-
An Unexpected Water Channel in the Light-Harvesting Complex of a Diatom: Implications for the Switch between Light Harvesting and Photoprotection.ACS Phys Chem Au. 2024 Aug 21;5(1):47-61. doi: 10.1021/acsphyschemau.4c00069. eCollection 2025 Jan 22. ACS Phys Chem Au. 2024. PMID: 39867443 Free PMC article.
-
Insights into substrate transport and water permeation in the mycobacterial transporter MmpL3.Biophys J. 2023 Jun 6;122(11):2342-2352. doi: 10.1016/j.bpj.2023.03.018. Epub 2023 Mar 16. Biophys J. 2023. PMID: 36926696 Free PMC article.
-
Quantitative insights into the mechanism of proton conduction and selectivity for the human voltage-gated proton channel Hv1.Proc Natl Acad Sci U S A. 2024 Sep 17;121(38):e2407479121. doi: 10.1073/pnas.2407479121. Epub 2024 Sep 11. Proc Natl Acad Sci U S A. 2024. PMID: 39259593 Free PMC article.
-
Ion channel structure and function of the MERS coronavirus E protein.Sci Adv. 2025 Jul 11;11(28):eadx1788. doi: 10.1126/sciadv.adx1788. Epub 2025 Jul 9. Sci Adv. 2025. PMID: 40632851 Free PMC article.
-
Efficient calculation of orientation-dependent lipid dynamics from membrane simulations.bioRxiv [Preprint]. 2024 Apr 15:2023.05.23.542012. doi: 10.1101/2023.05.23.542012. bioRxiv. 2024. PMID: 37292992 Free PMC article. Preprint.
References
References and notes:
References for Materials and Methods
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
