

Varicella-zoster virus proteome-wide T-cell screening demonstrates low prevalence of virus-specific CD8 T-cells in latently infected human trigeminal ganglia
- PMID: 37308917
- PMCID: PMC10259006
- DOI: 10.1186/s12974-023-02820-y
Varicella-zoster virus proteome-wide T-cell screening demonstrates low prevalence of virus-specific CD8 T-cells in latently infected human trigeminal ganglia
Abstract
Background: Trigeminal ganglia (TG) neurons are an important site of lifelong latent varicella-zoster virus (VZV) infection. Although VZV-specific T-cells are considered pivotal to control virus reactivation, their protective role at the site of latency remains uncharacterized.
Methods: Paired blood and TG specimens were obtained from ten latent VZV-infected adults, of which nine were co-infected with herpes simplex virus type 1 (HSV-1). Short-term TG-derived T-cell lines (TG-TCL), generated by mitogenic stimulation of TG-derived T-cells, were probed for HSV-1- and VZV-specific T-cells using flow cytometry. We also performed VZV proteome-wide screening of TG-TCL to determine the fine antigenic specificity of VZV reactive T-cells. Finally, the relationship between T-cells and latent HSV-1 and VZV infections in TG was analyzed by reverse transcription quantitative PCR (RT-qPCR) and in situ analysis for T-cell proteins and latent viral transcripts.
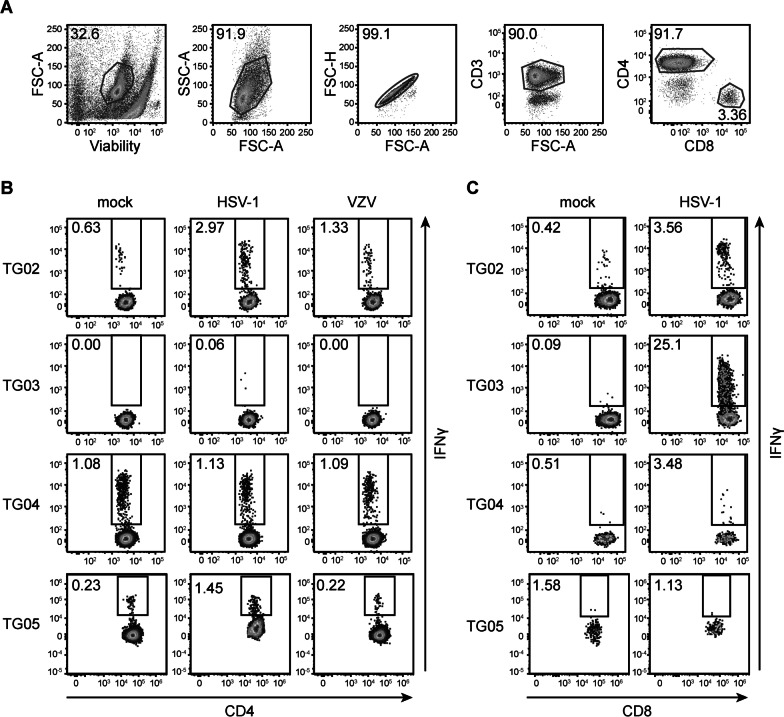
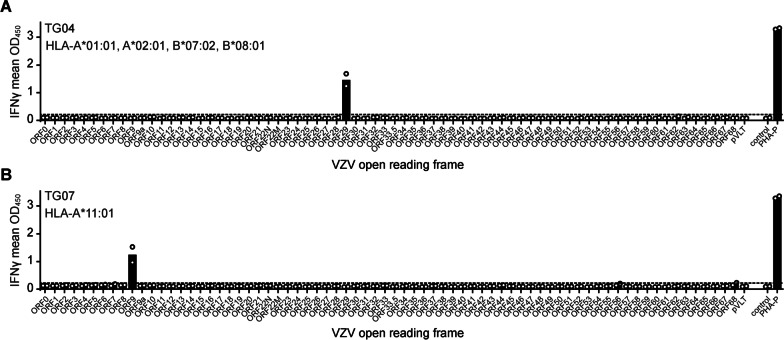
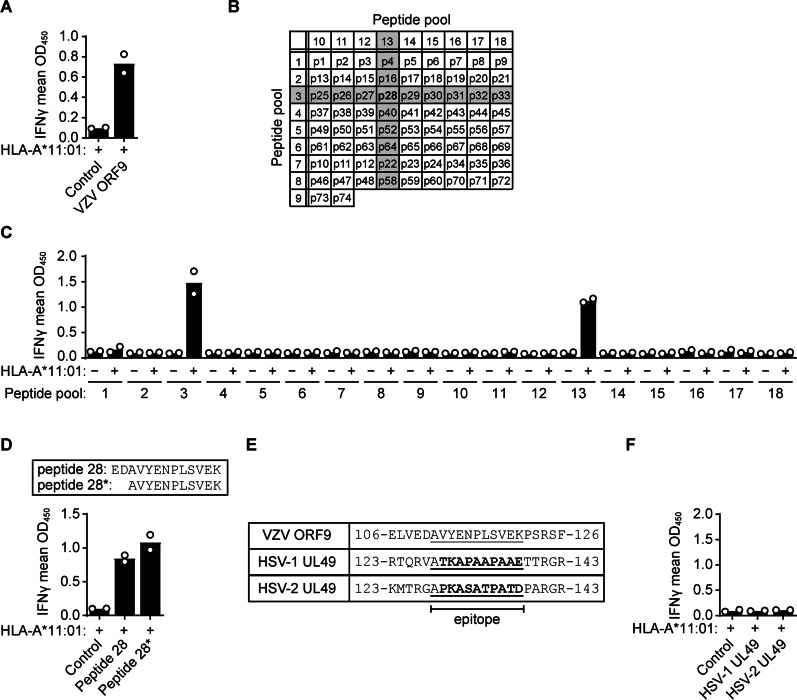
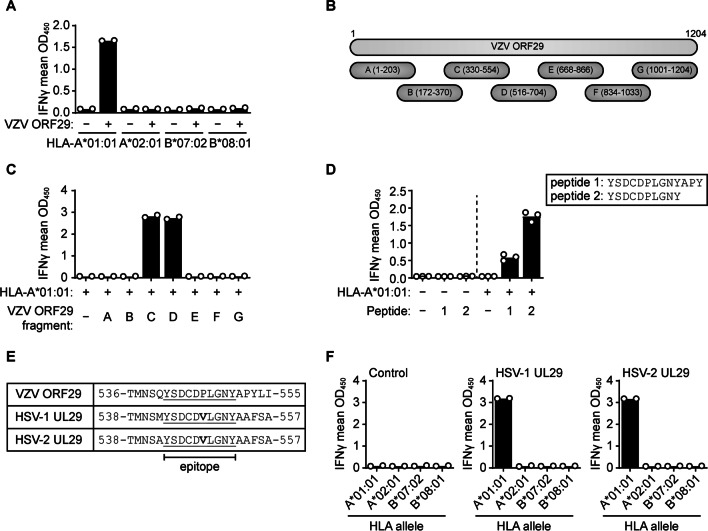
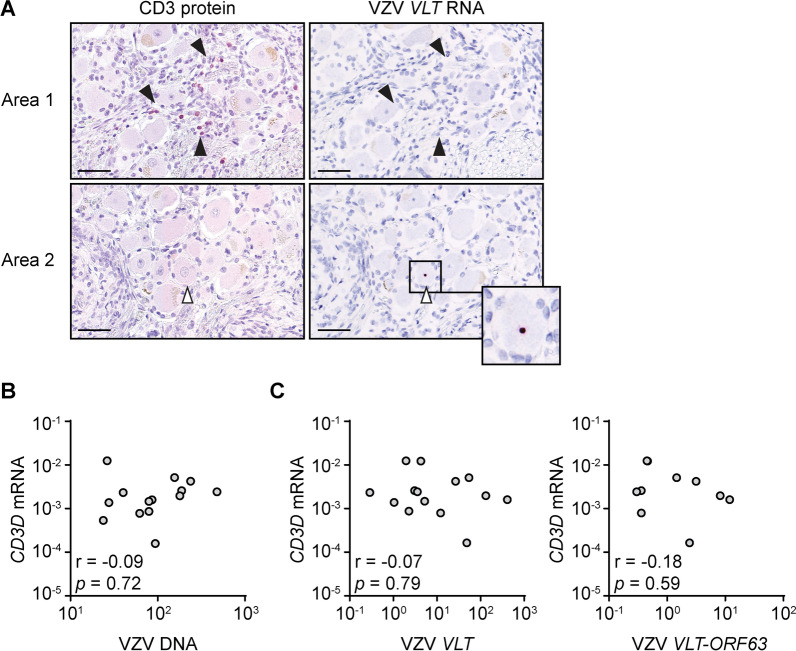
Results: VZV proteome-wide analysis of ten TG-TCL identified two VZV antigens recognized by CD8 T-cells in two separate subjects. The first was an HSV-1/VZV cross-reactive CD8 T-cell epitope, whereas the second TG harbored CD8 T-cells reactive with VZV specifically and not the homologous peptide in HSV-1. In silico analysis showed that HSV-1/VZV cross reactivity of TG-derived CD8 T-cells reactive with ten previously identified HSV-1 epitopes was unlikely, suggesting that HSV-1/VZV cross-reactive T-cells are not a common feature in dually infected TG. Finally, no association was detected between T-cell infiltration and VZV latency transcript abundance in TG by RT-qPCR or in situ analyses.
Conclusions: The low presence of VZV- compared to HSV-1-specific CD8 T-cells in human TG suggests that VZV reactive CD8 T-cells play a limited role in maintaining VZV latency.
Keywords: Herpes simplex virus; Human; Latency; T-cells; Trigeminal ganglion; Varicella-zoster virus.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Knipe DMPMH, editor. Field’s Virology. 7. Philadelphia: Lipincott Williams & Wilkins; 2020.
-
- Ouwendijk WJD, Depledge DP, Rajbhandari L, Lenac Rovis T, Jonjic S, Breuer J, Venkatesan A, Verjans GMGM, Sadaoka T. Varicella-zoster virus VLT-ORF63 fusion transcript induces broad viral gene expression during reactivation from neuronal latency. Nat Commun. 2020;11:6324. doi: 10.1038/s41467-020-20031-4. - DOI - PMC - PubMed
-
- Zerboni L, Sobel RA, Lai M, Triglia R, Steain M, Abendroth A, Arvin A. Apparent expression of varicella-zoster virus proteins in latency resulting from reactivity of murine and rabbit antibodies with human blood group a determinants in sensory neurons. J Virol. 2012;86:578–583. doi: 10.1128/JVI.05950-11. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
