

BMPER is a marker of adipose progenitors and adipocytes and a positive modulator of adipogenesis
- PMID: 37311809
- PMCID: PMC10264349
- DOI: 10.1038/s42003-023-05011-w
BMPER is a marker of adipose progenitors and adipocytes and a positive modulator of adipogenesis
Abstract
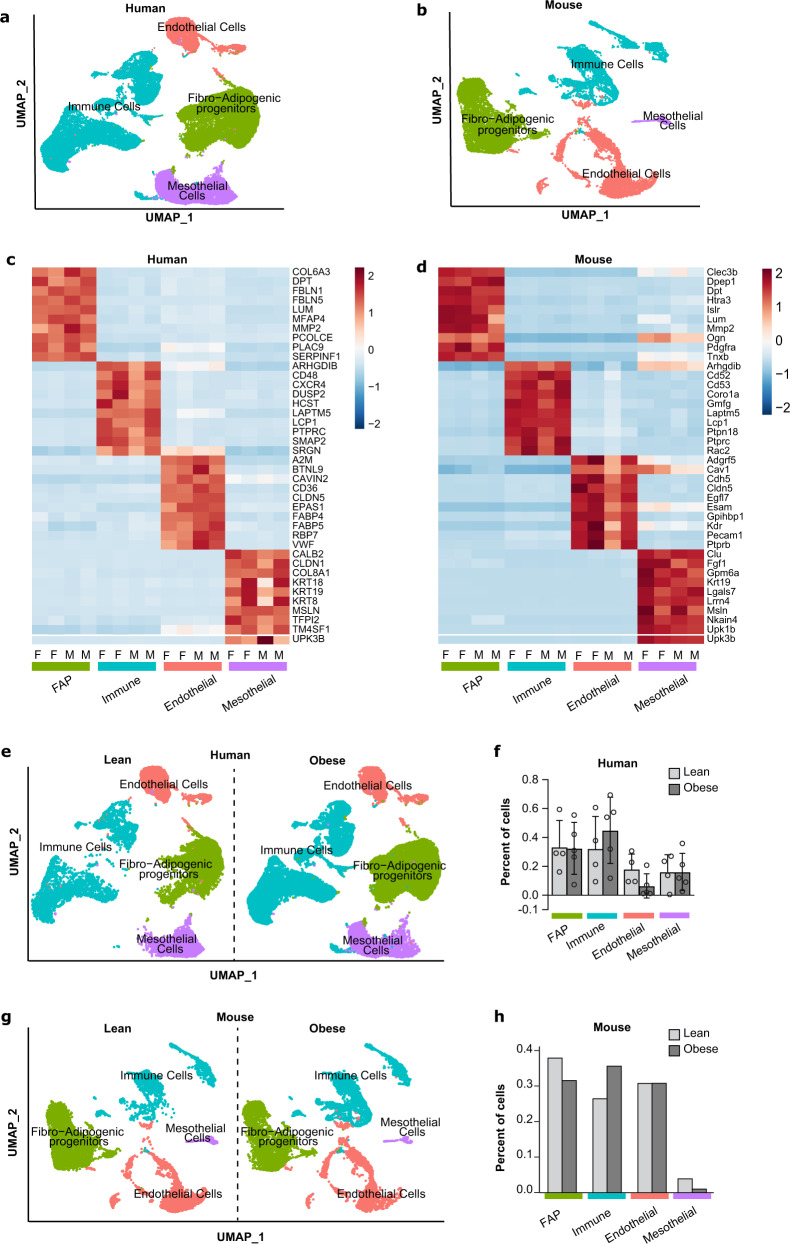
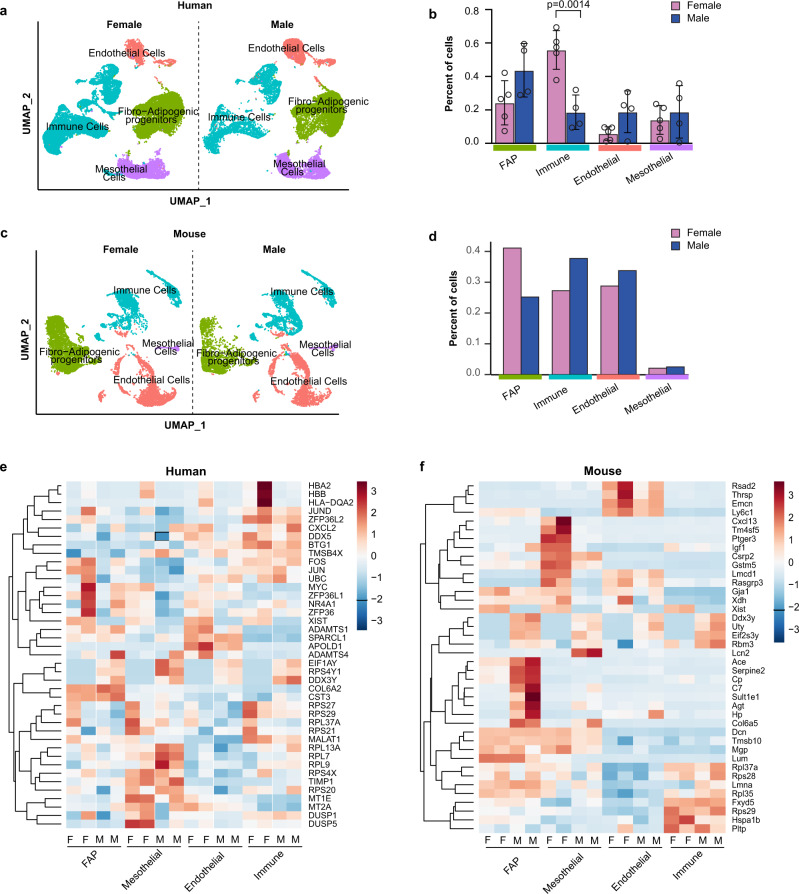
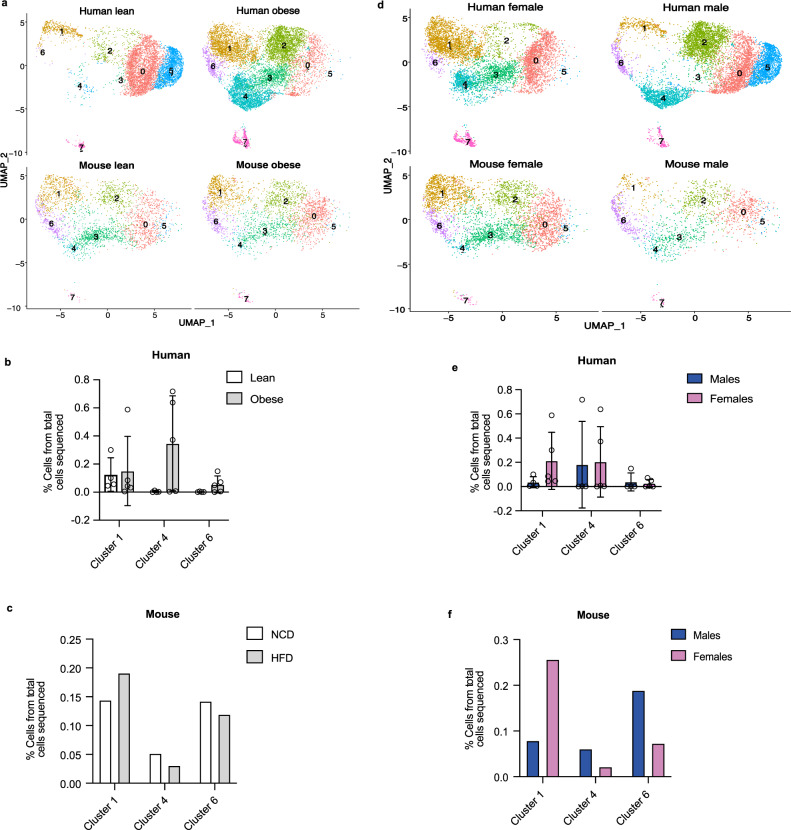
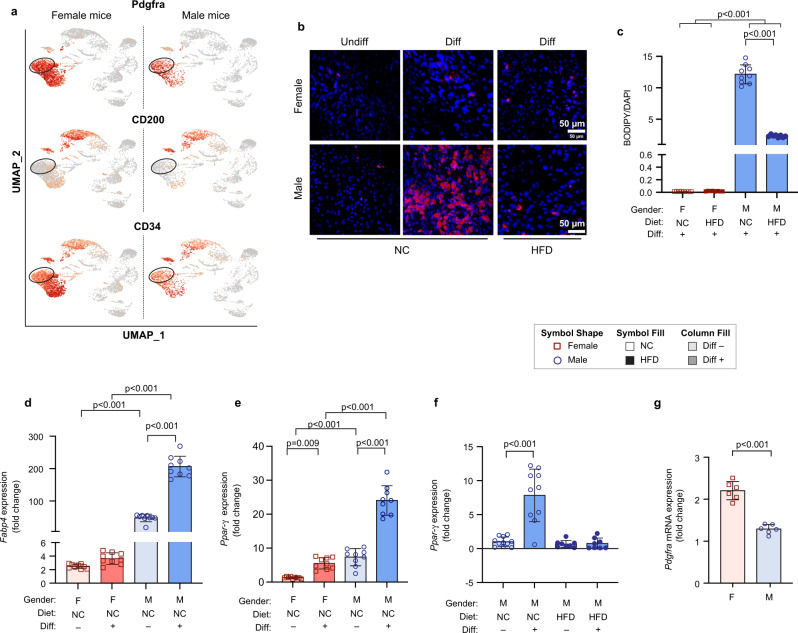
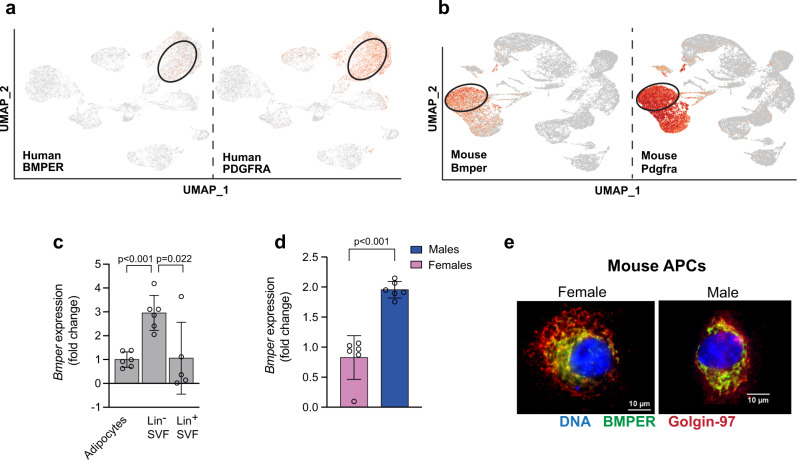
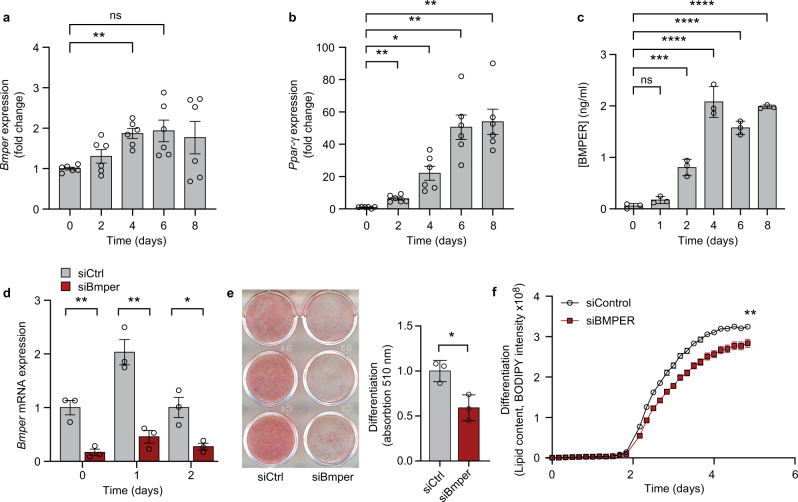
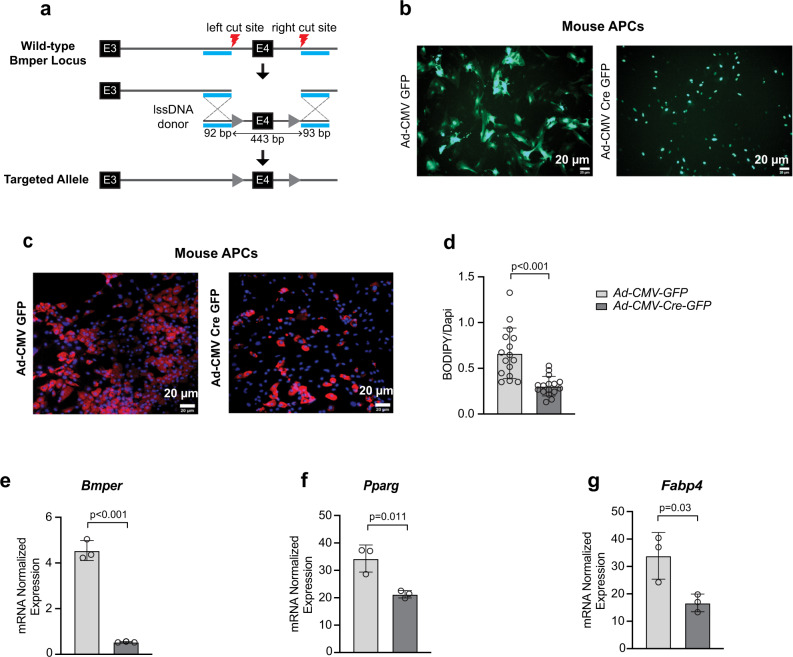
Autocrine and paracrine signaling regulating adipogenesis in white adipose tissue remains largely unclear. Here we used single-cell RNA-sequencing (RNA-seq) and single nuclei RNA-sequencing (snRNA-seq) to identify markers of adipose progenitor cells (APCs) and adipogenic modulators in visceral adipose tissue (VAT) of humans and mice. Our study confirmed the presence of major cellular clusters in humans and mice and established important sex and diet-specific dissimilarities in cell proportions. Here we show that bone morphogenetic protein (BMP)-binding endothelial regulator (BMPER) is a conserved marker for APCs and adipocytes in VAT in humans and mice. Further, BMPER is highly enriched in lineage negative stromal vascular cells and its expression is significantly higher in visceral compared to subcutaneous APCs in mice. BMPER expression and release peaked by day four post-differentiation in 3T3-L1 preadipocytes. We reveal that BMPER is required for adipogenesis both in 3T3-L1 preadipocytes and in mouse APCs. Together, this study identified BMPER as a positive modulator of adipogenesis.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
