

Canonical BAF complex activity shapes the enhancer landscape that licenses CD8+ T cell effector and memory fates
- PMID: 37315534
- PMCID: PMC10281564
- DOI: 10.1016/j.immuni.2023.05.005
Canonical BAF complex activity shapes the enhancer landscape that licenses CD8+ T cell effector and memory fates
Abstract
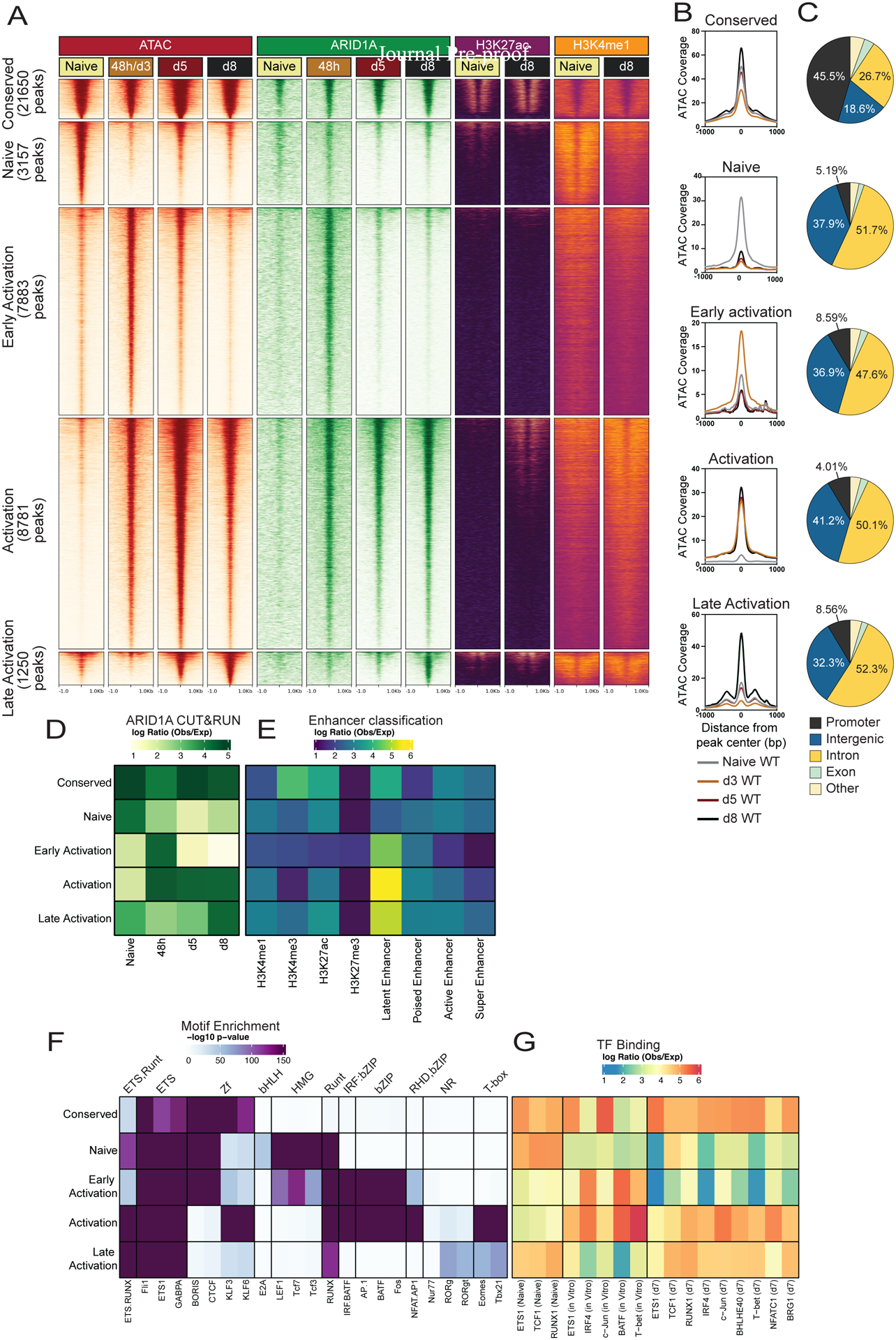
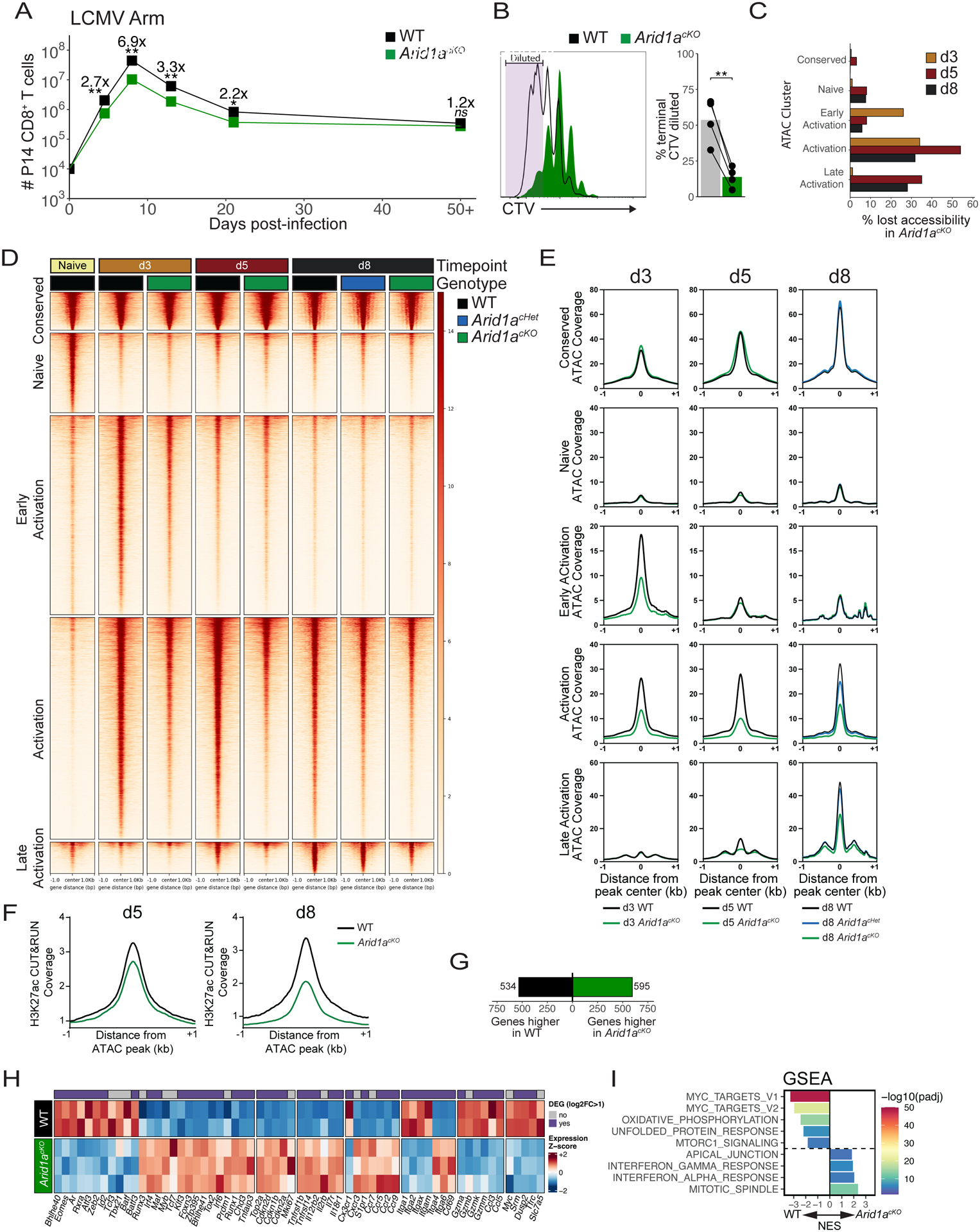
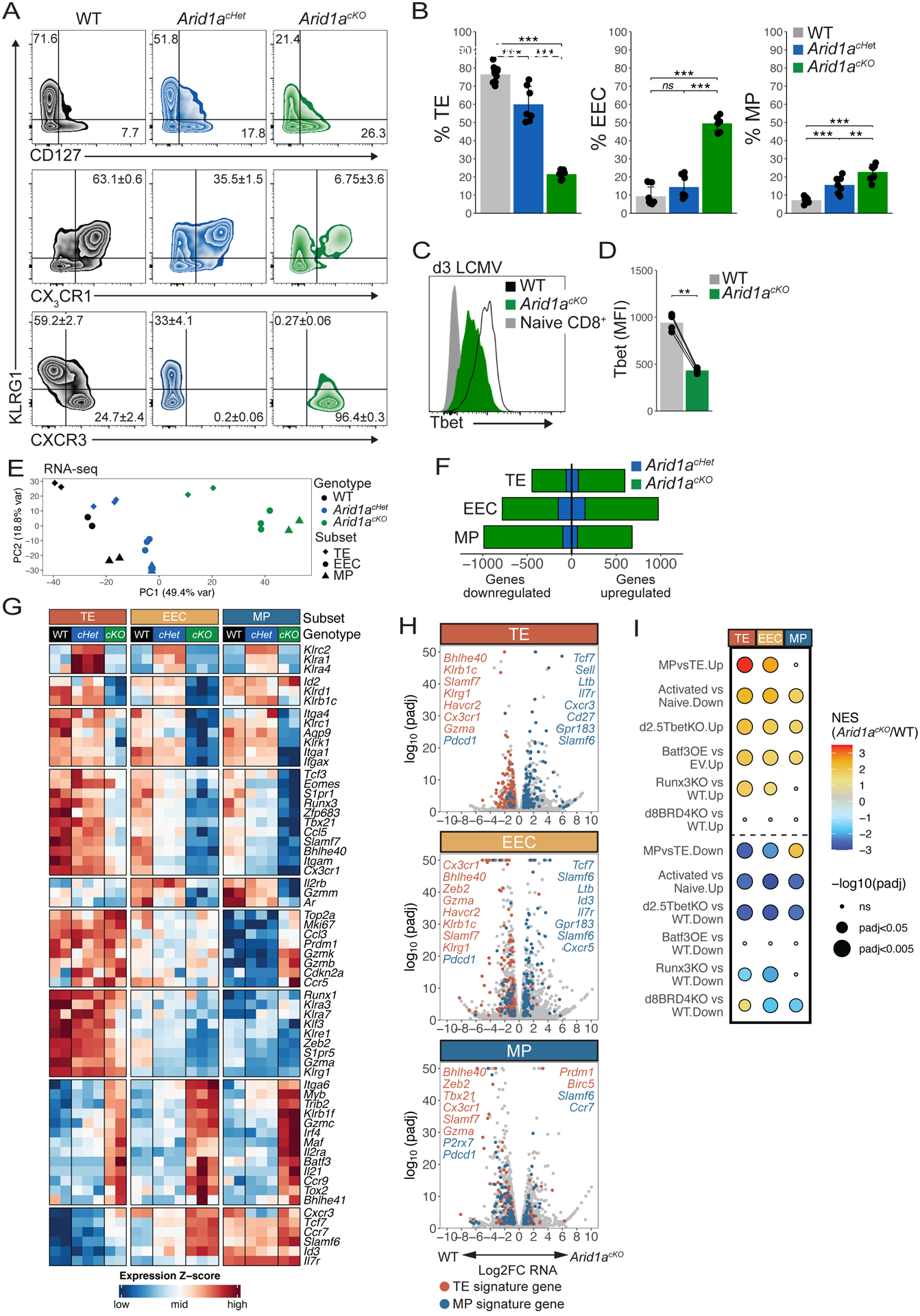
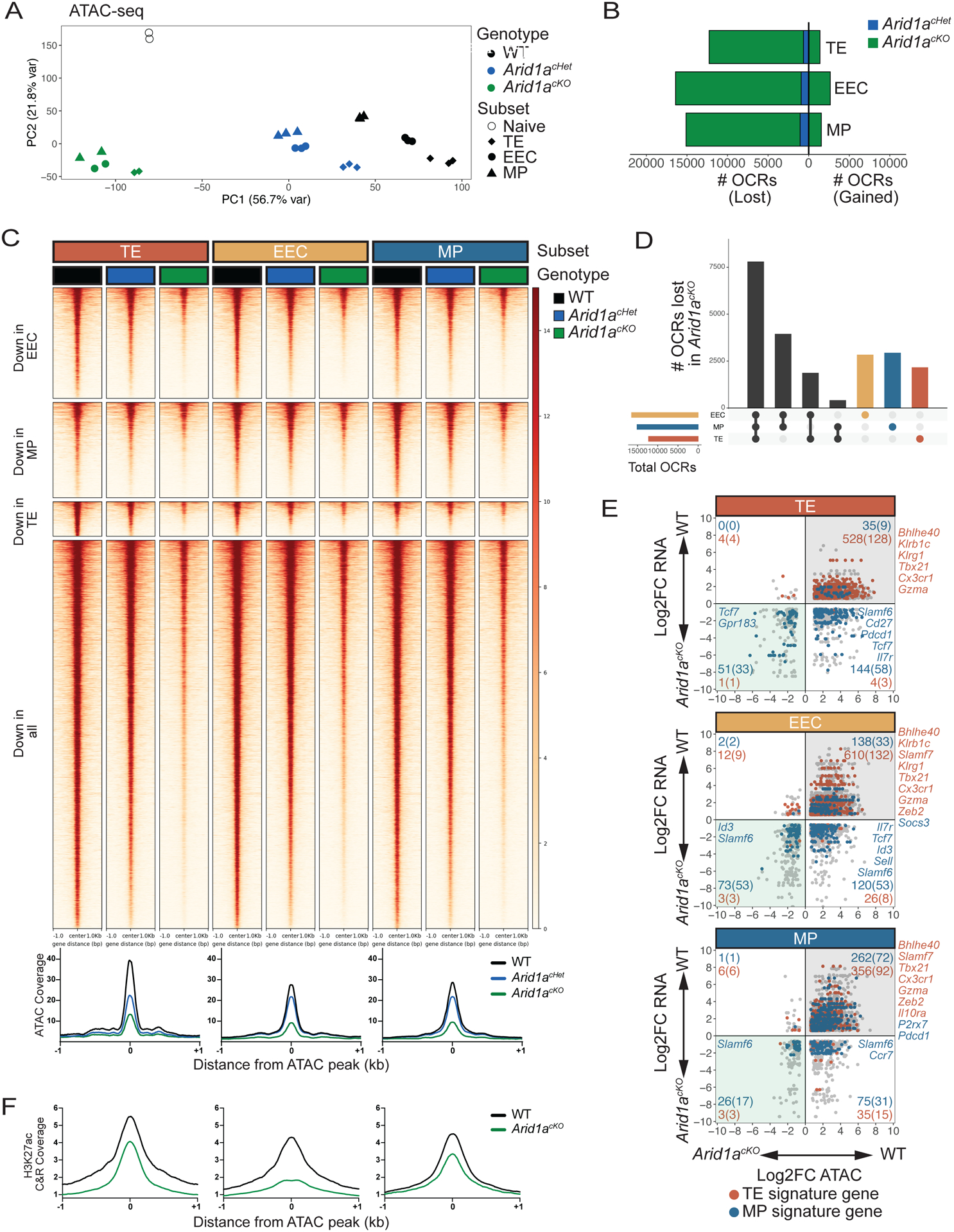
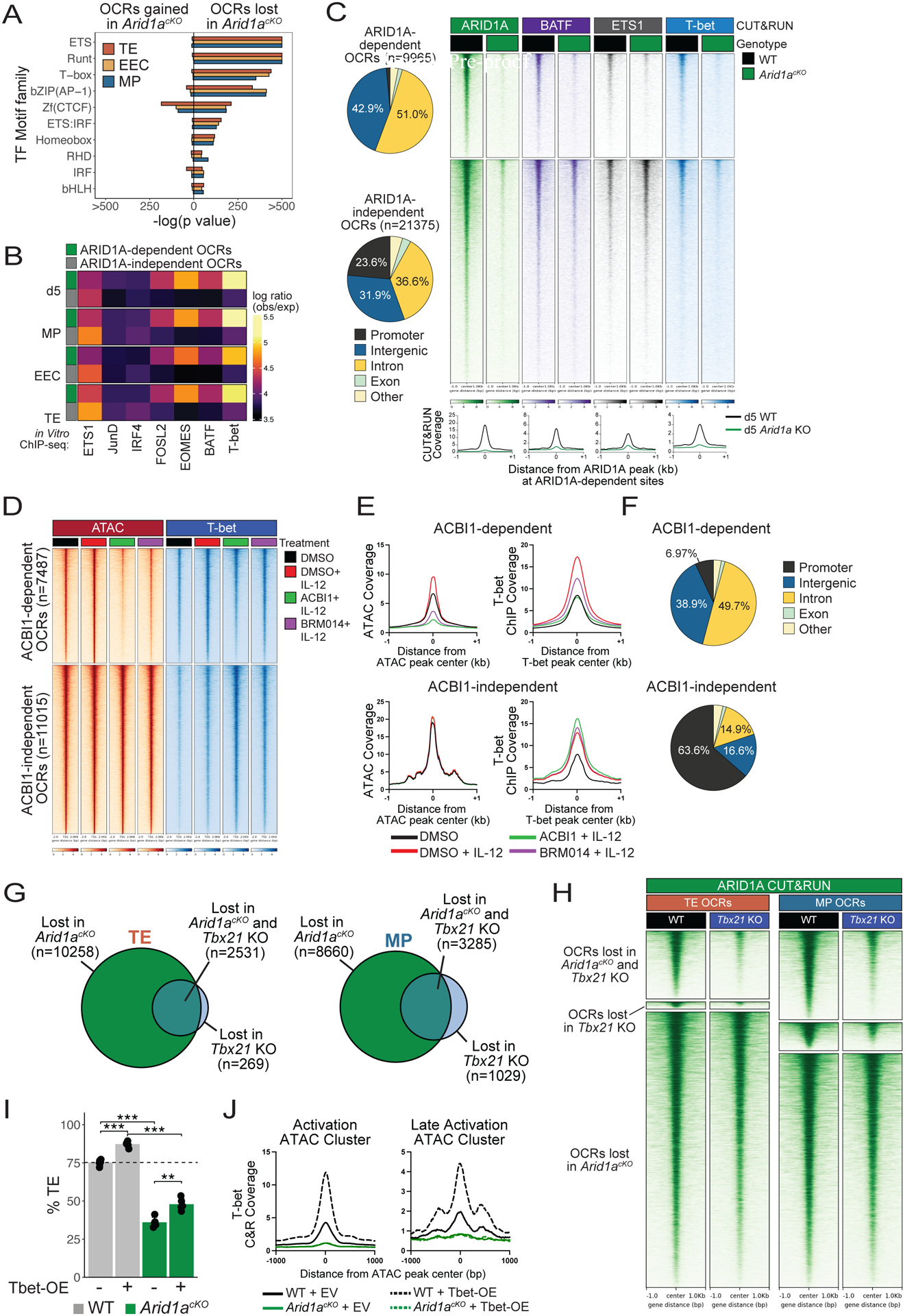
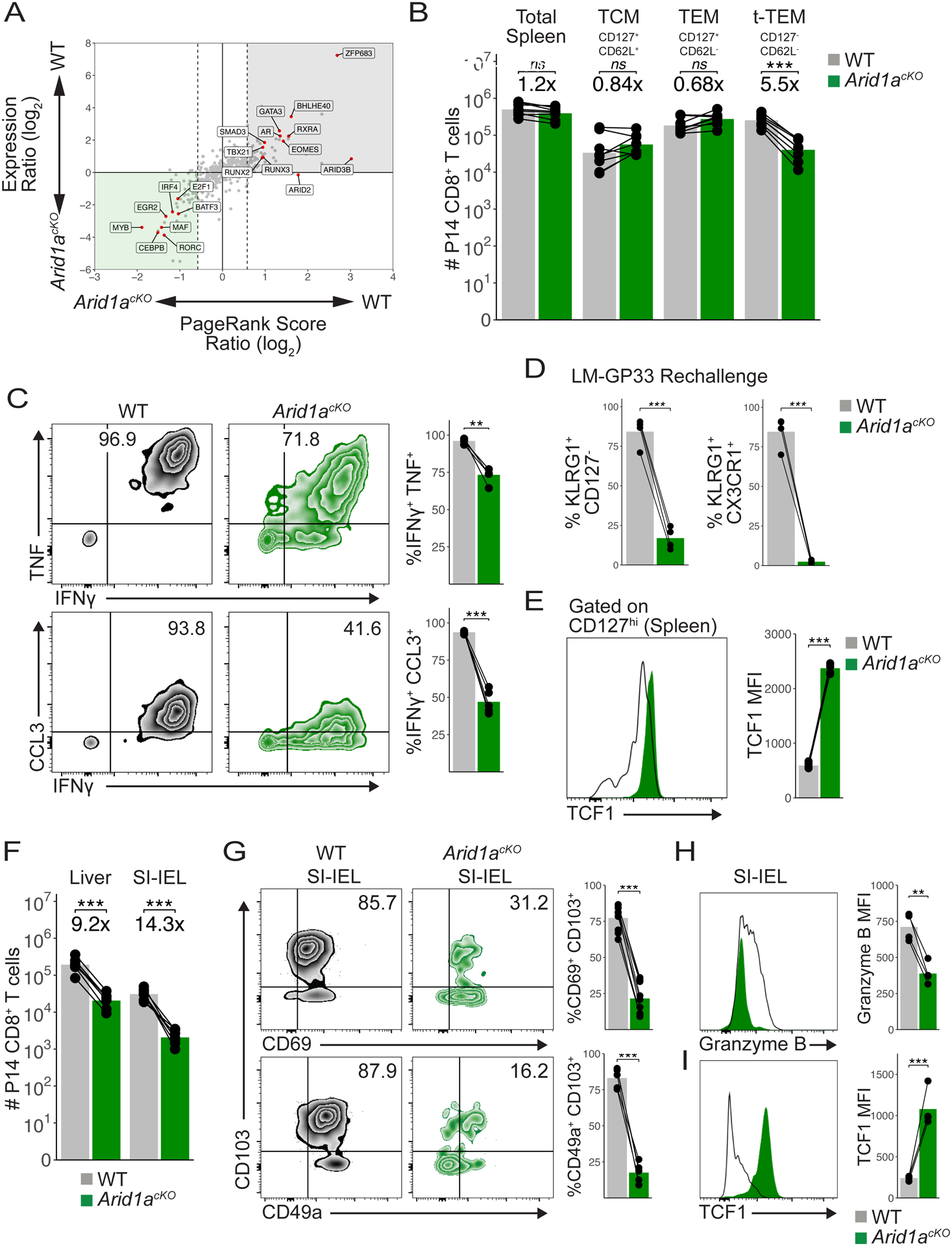
CD8+ T cells provide host protection against pathogens by differentiating into distinct effector and memory cell subsets, but how chromatin is site-specifically remodeled during their differentiation is unclear. Due to its critical role in regulating chromatin and enhancer accessibility through its nucleosome remodeling activities, we investigated the role of the canonical BAF (cBAF) chromatin remodeling complex in antiviral CD8+ T cells during infection. ARID1A, a subunit of cBAF, was recruited early after activation and established de novo open chromatin regions (OCRs) at enhancers. Arid1a deficiency impaired the opening of thousands of activation-induced enhancers, leading to loss of TF binding, dysregulated proliferation and gene expression, and failure to undergo terminal effector differentiation. Although Arid1a was dispensable for circulating memory cell formation, tissue-resident memory (Trm) formation was strongly impaired. Thus, cBAF governs the enhancer landscape of activated CD8+ T cells that orchestrates TF recruitment and activity and the acquisition of specific effector and memory differentiation states.
Keywords: ARID1A; BAF complex; CD8(+) T cells; antiviral immunity; chromatin remodeling; effector T cell; epigenetics; immunotherapy; memory T cell.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.M.K. is on the scientific advisory boards and has equity in EvolveImmune Therapeutics, Affini-T Therapeutics, Arvinas, and Pfizer.
Figures
References
-
- Gerlach C, Moseman EA, Loughhead SM, Alvarez D, Zwijnenburg AJ, Waanders L, Garg R, de la Torre JC, and von Andrian UH (2016). The Chemokine Receptor CX3CR1 Defines Three Antigen-Experienced CD8 T Cell Subsets with Distinct Roles in Immune Surveillance and Homeostasis. Immunity 45, 1270–1284. 10.1016/j.immuni.2016.10.018. - DOI - PMC - PubMed
-
- Herndler-Brandstetter D, Ishigame H, Shinnakasu R, Plajer V, Stecher C, Zhao J, Lietzenmayer M, Kroehling L, Takumi A, Kometani K, et al. (2018). KLRG1+ Effector CD8+ T Cells Lose KLRG1, Differentiate into All Memory T Cell Lineages, and Convey Enhanced Protective Immunity. Immunity 48, 716–729.e8. 10.1016/j.immuni.2018.03.015. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
