

Fibroblast-derived PI16 sustains inflammatory pain via regulation of CD206+ myeloid cells
- PMID: 37315702
- PMCID: PMC10527931
- DOI: 10.1016/j.bbi.2023.06.011
Fibroblast-derived PI16 sustains inflammatory pain via regulation of CD206+ myeloid cells
Abstract
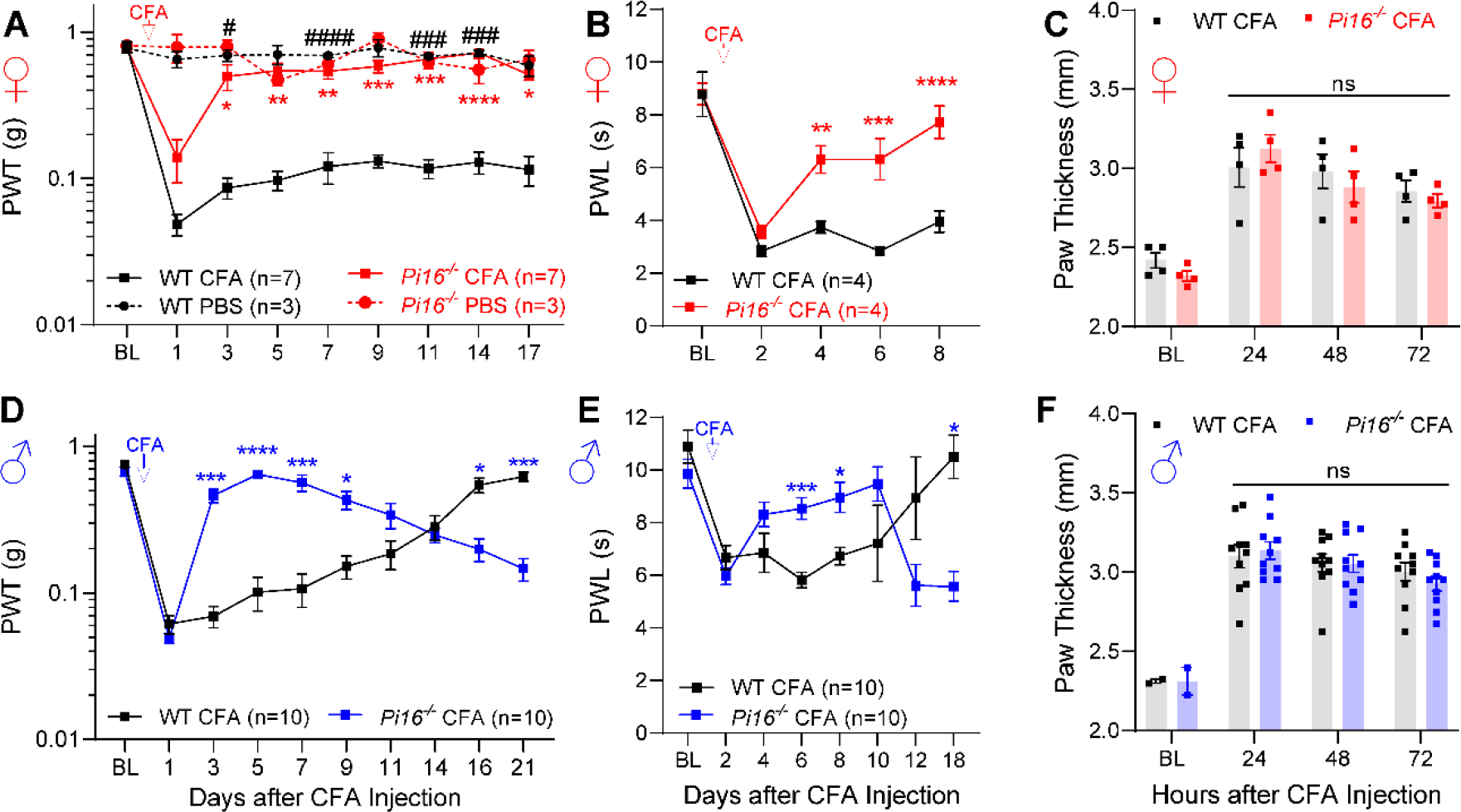
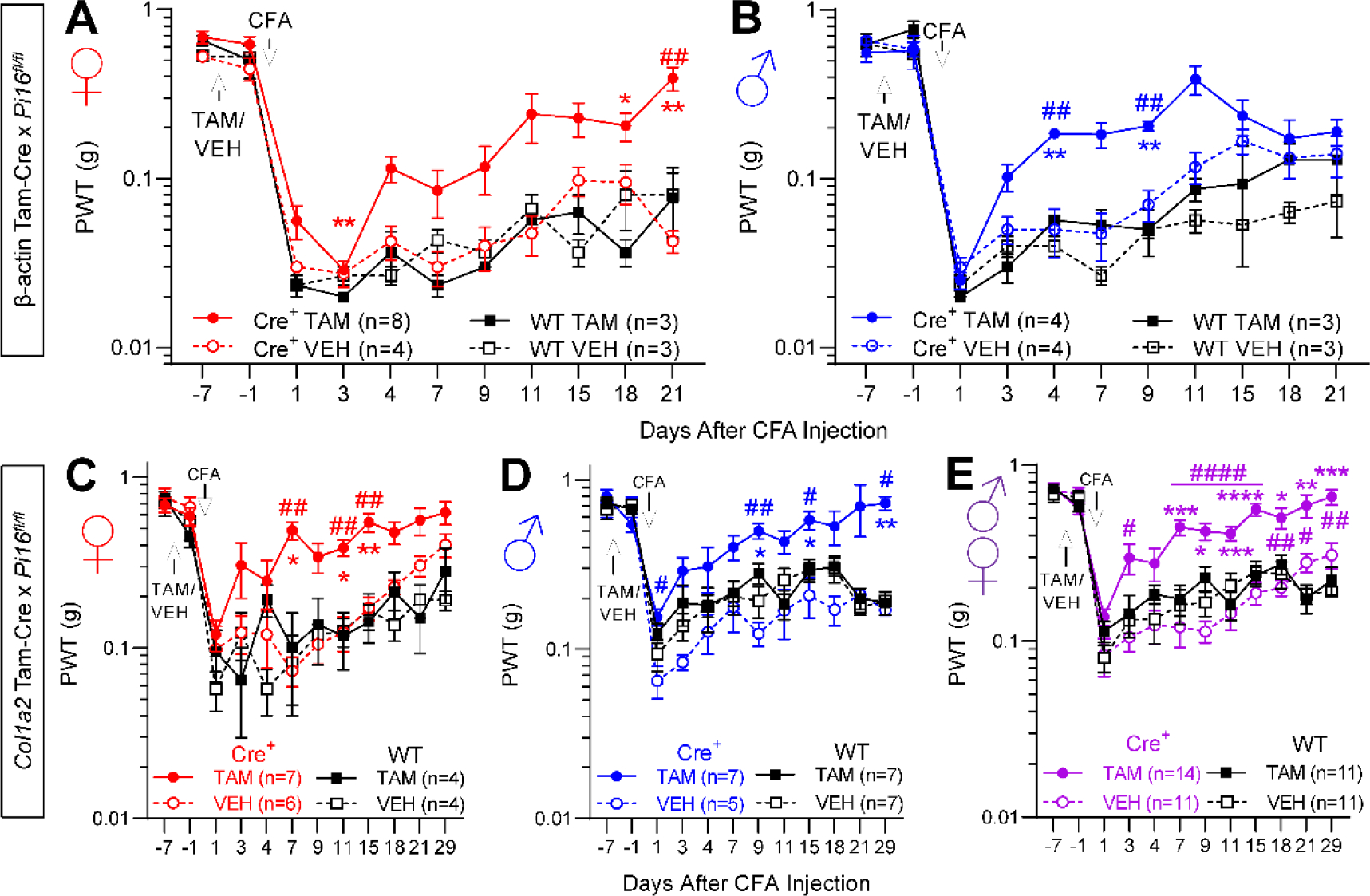
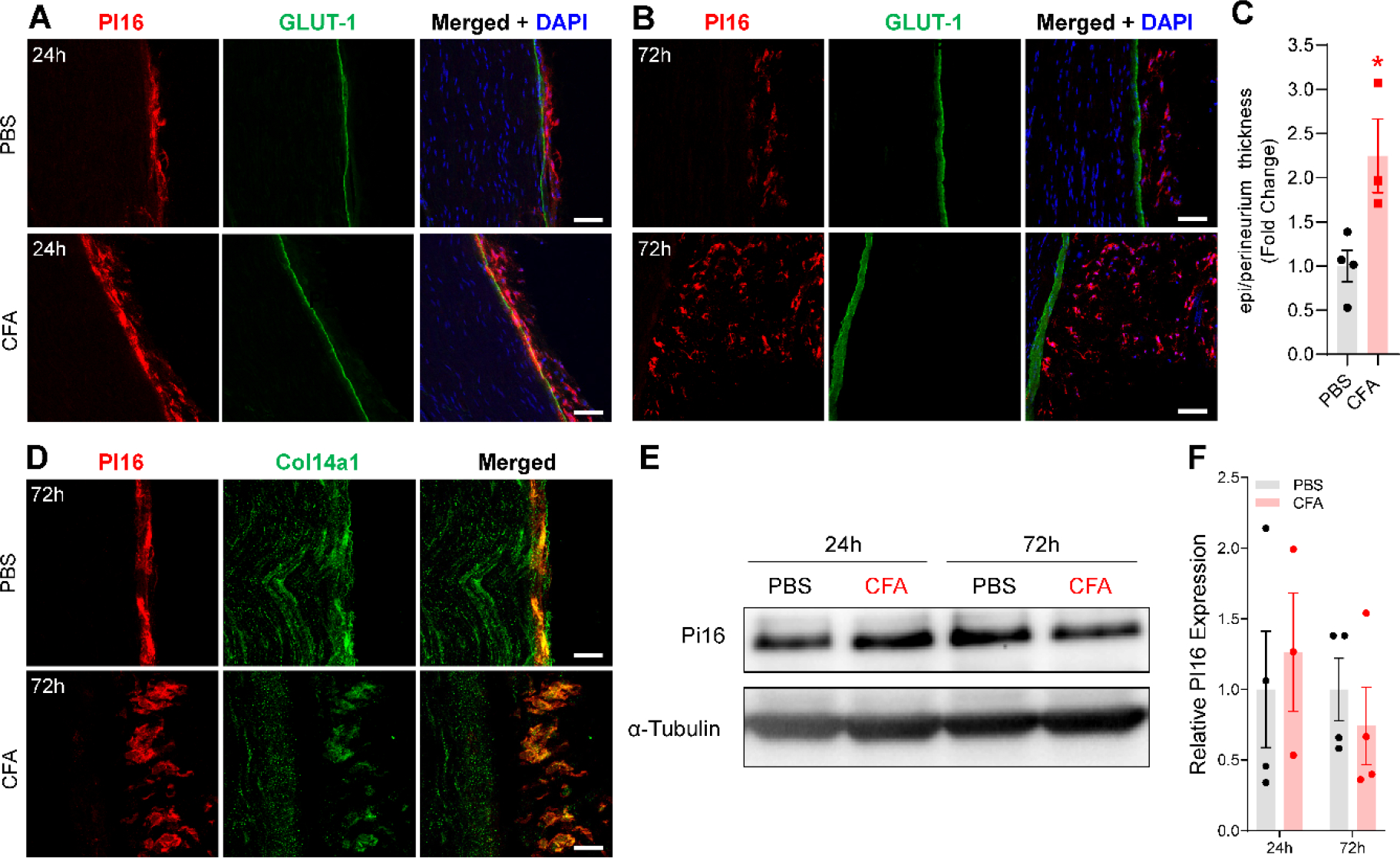
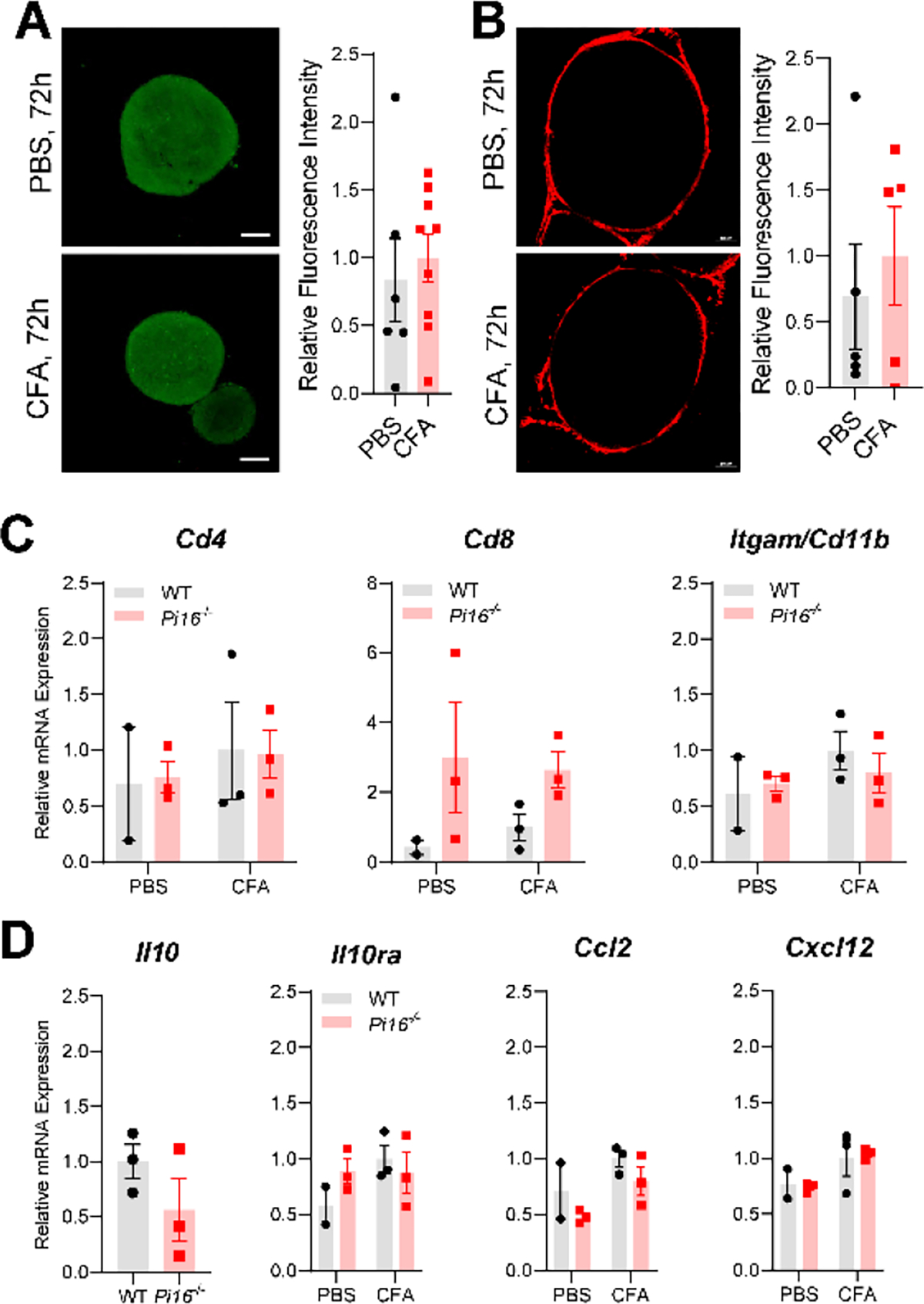
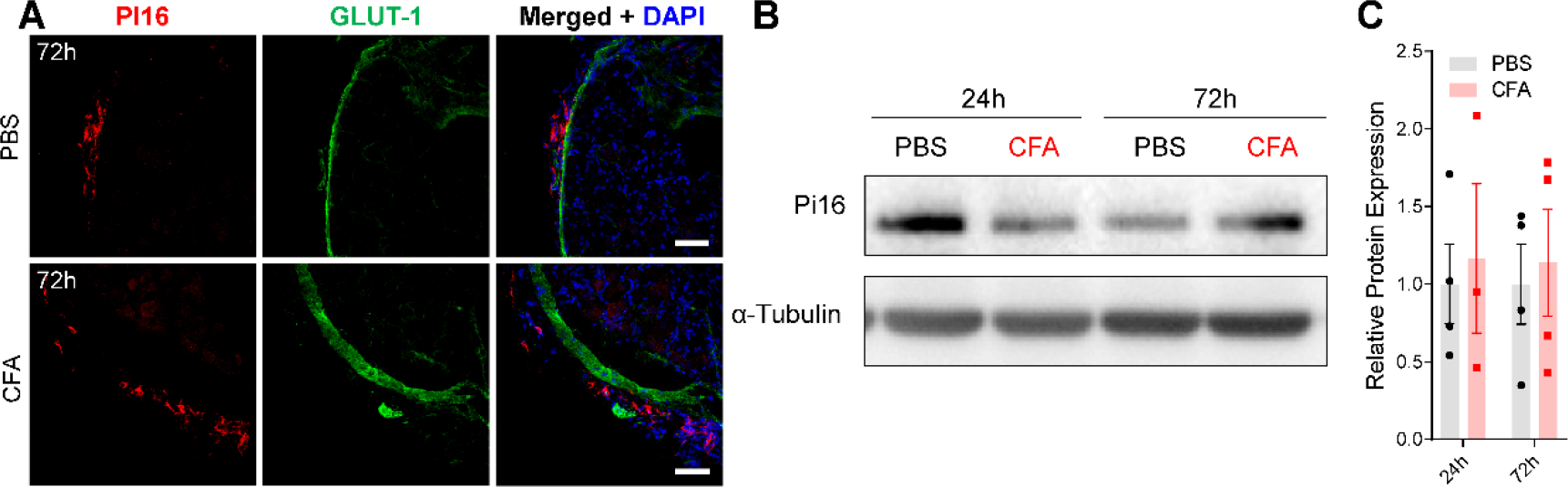
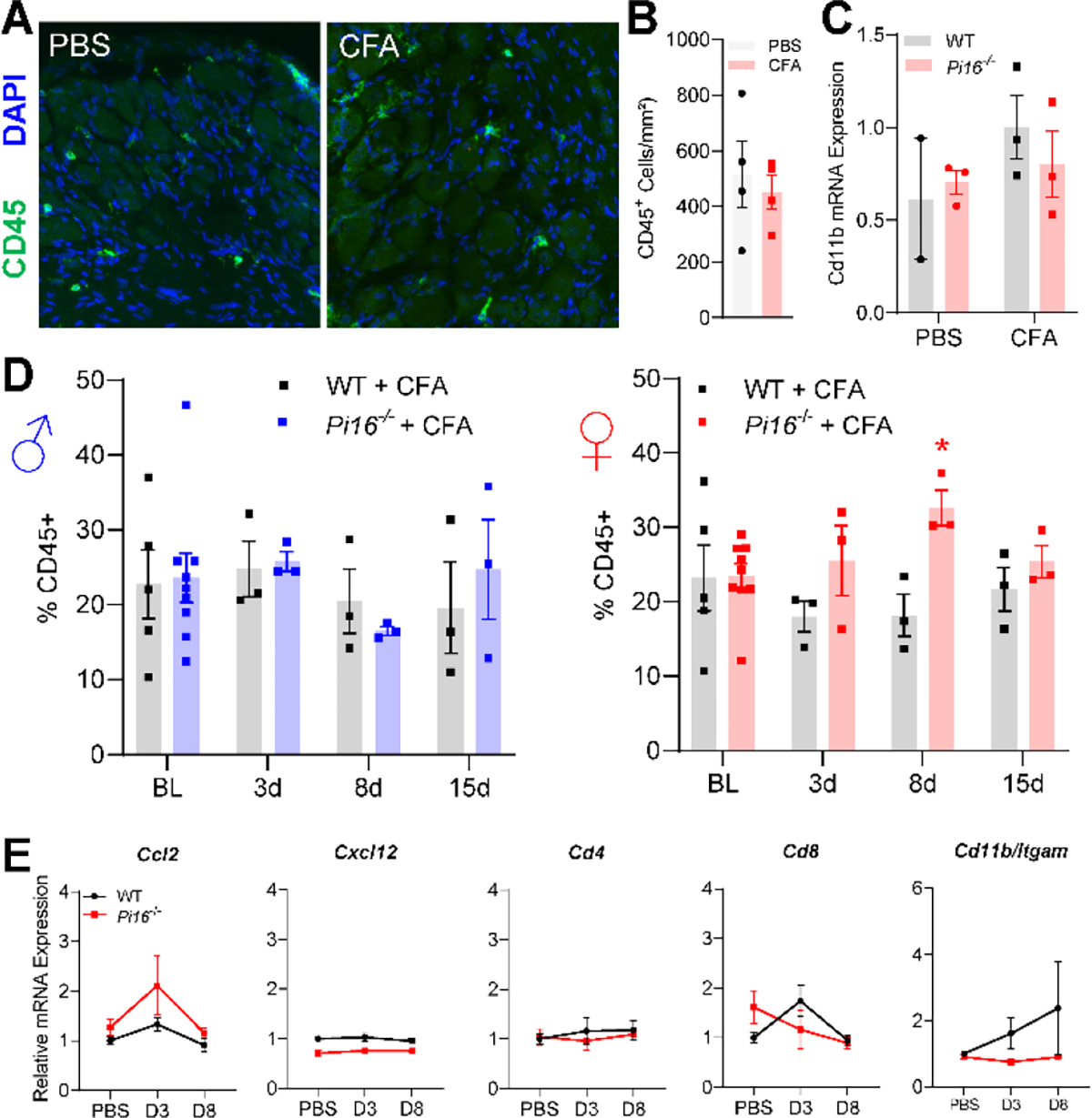
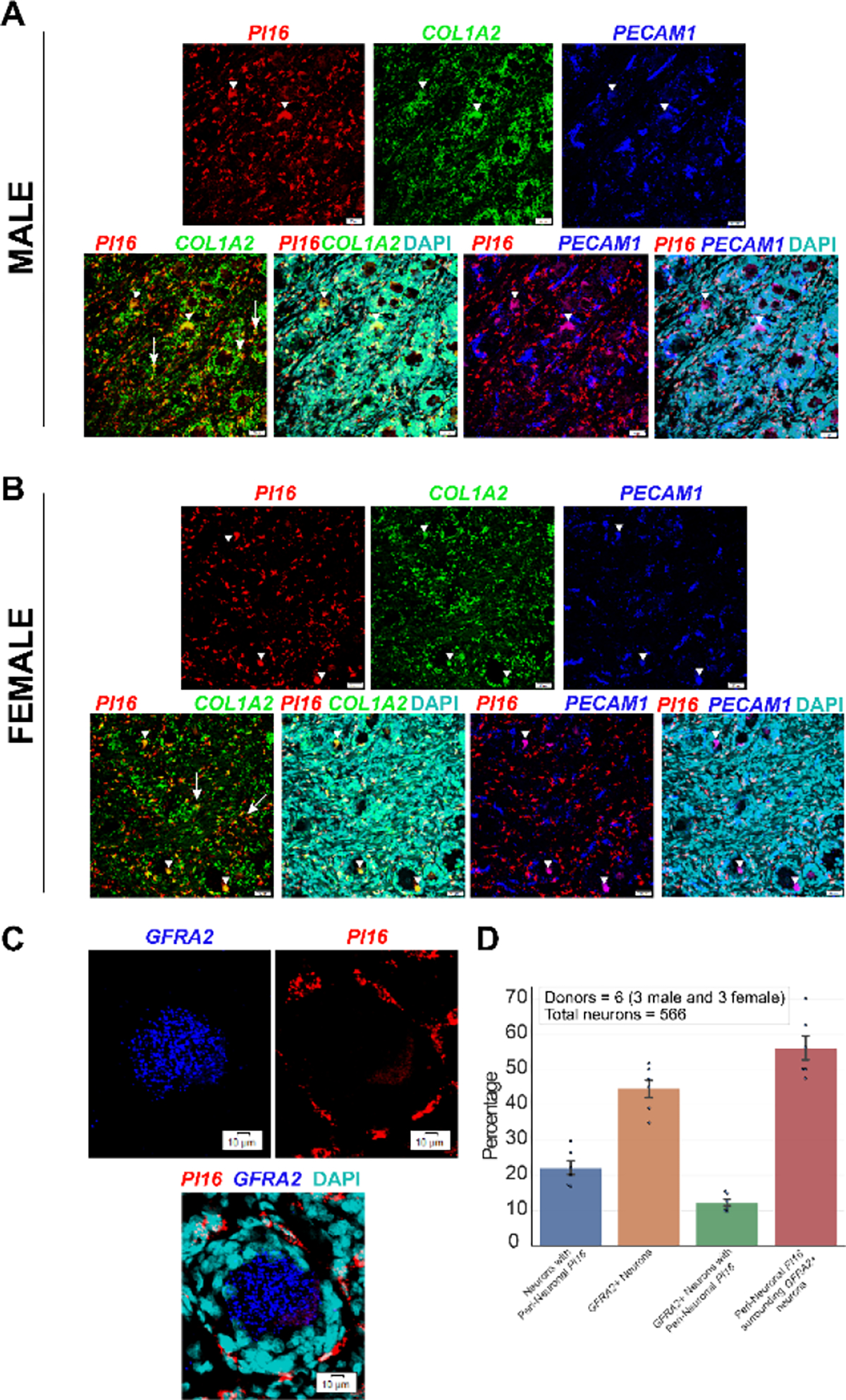
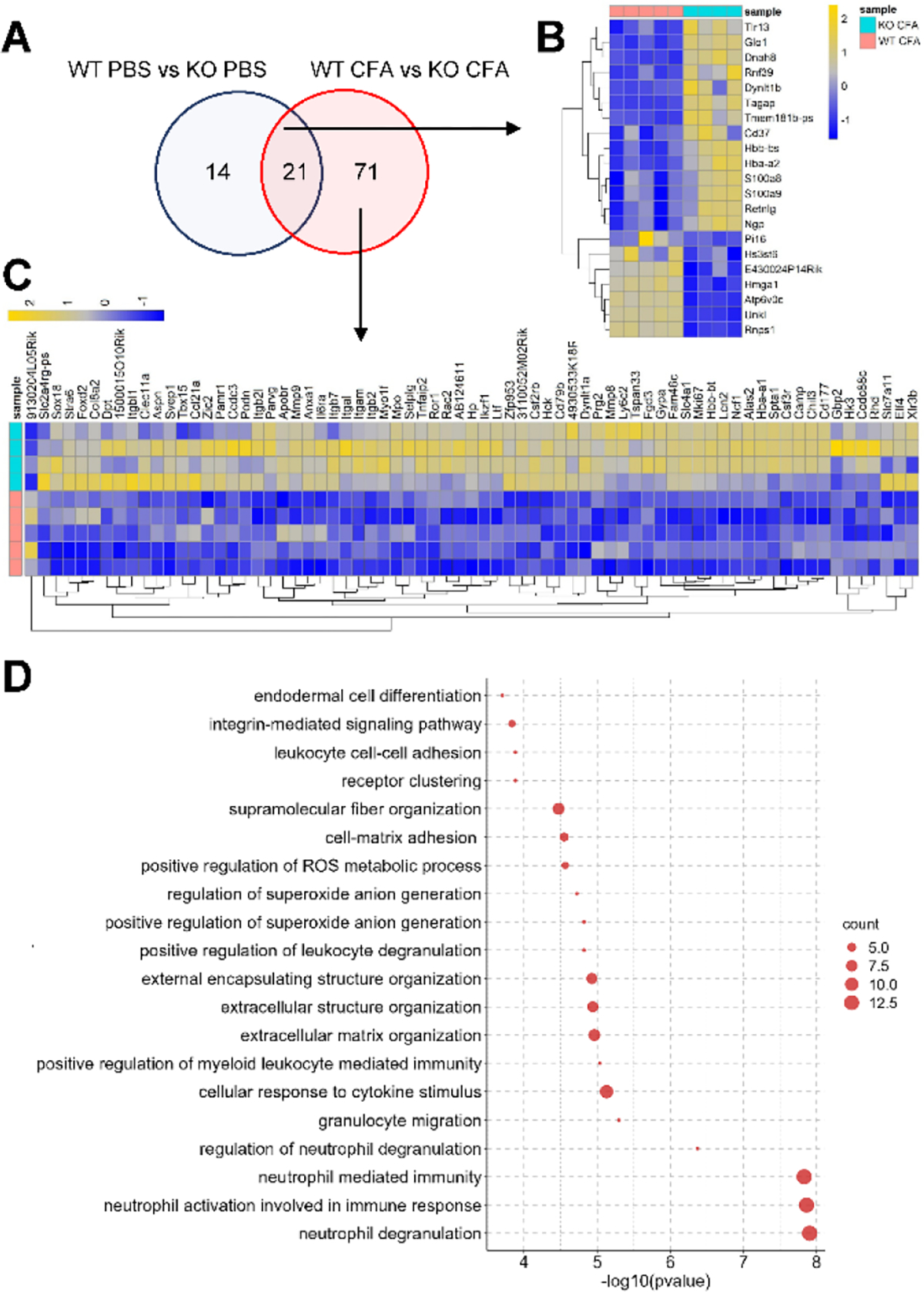
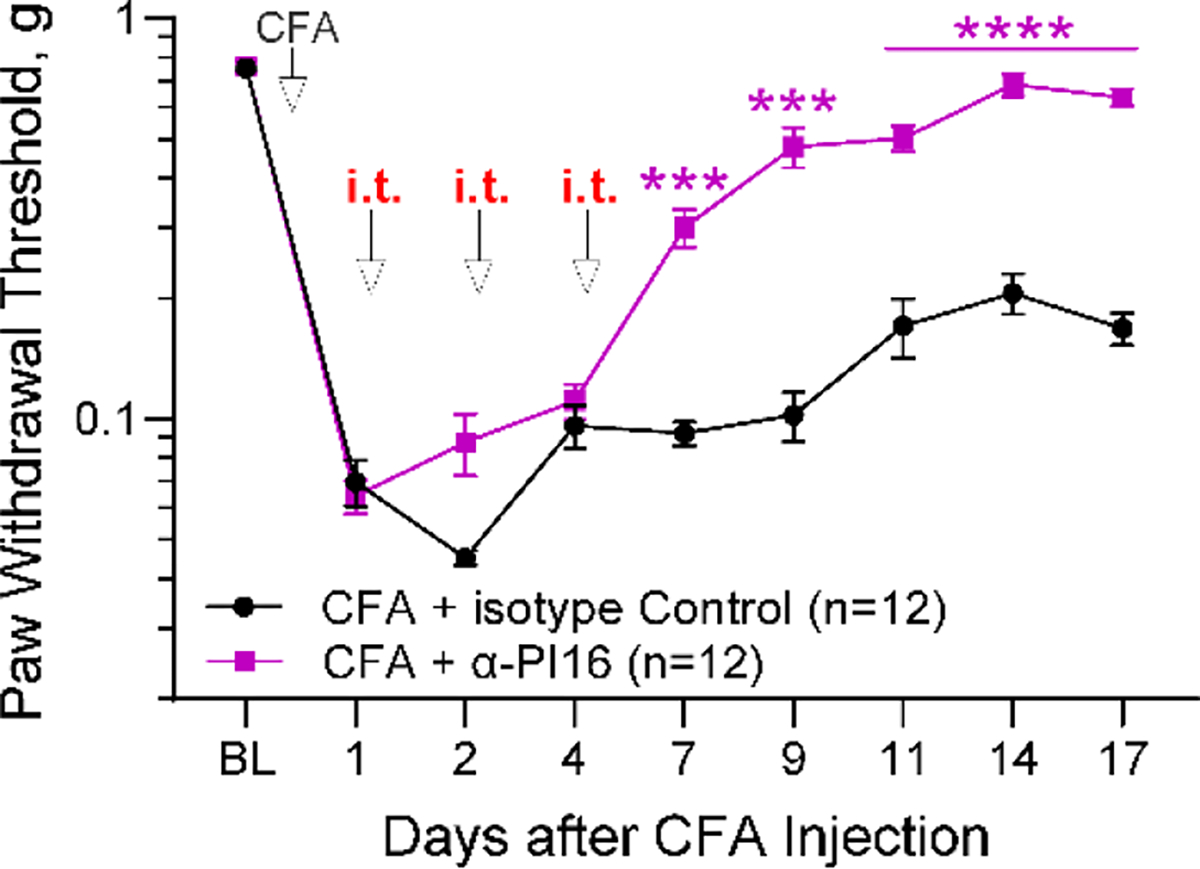
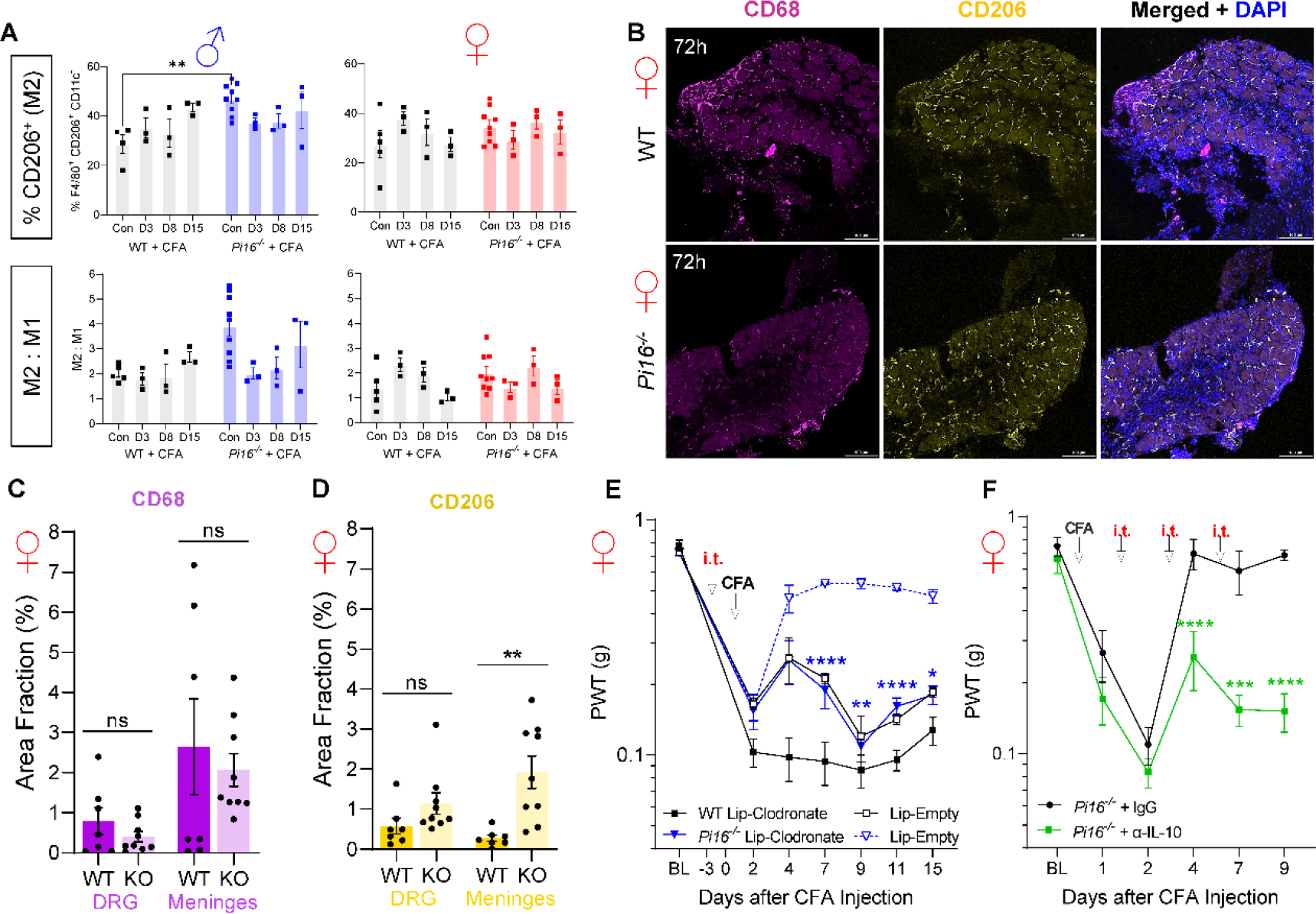
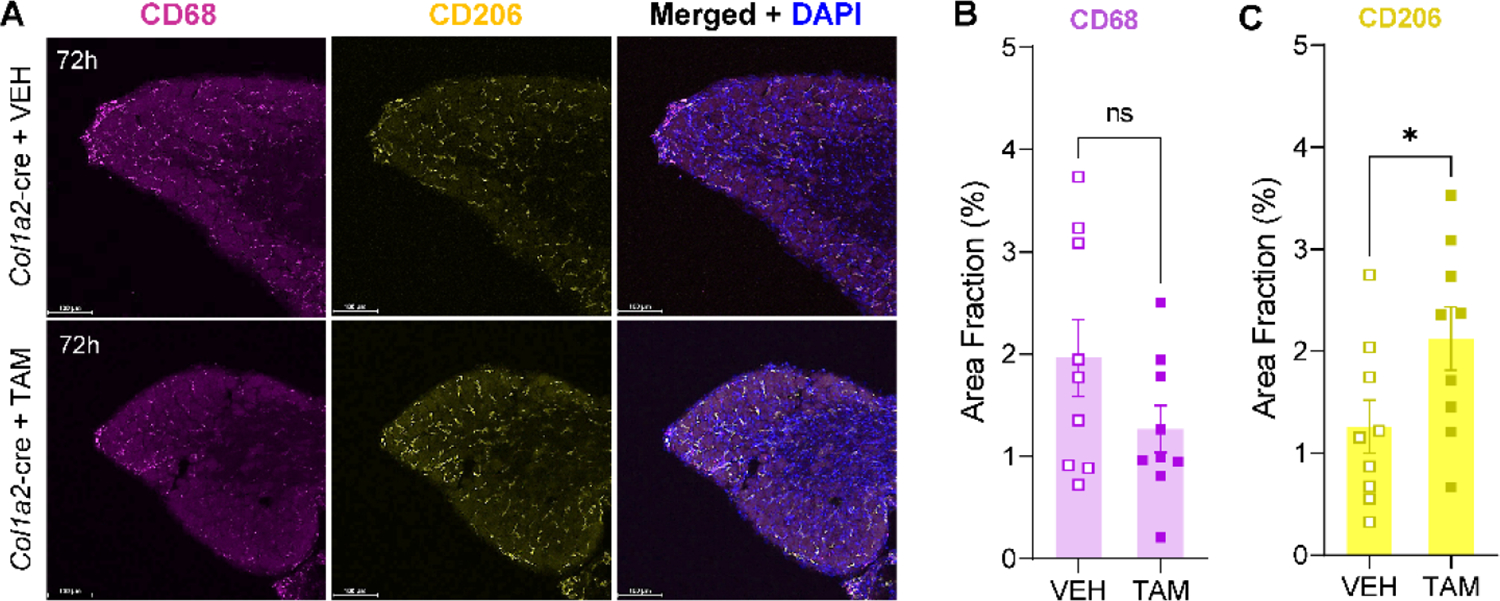
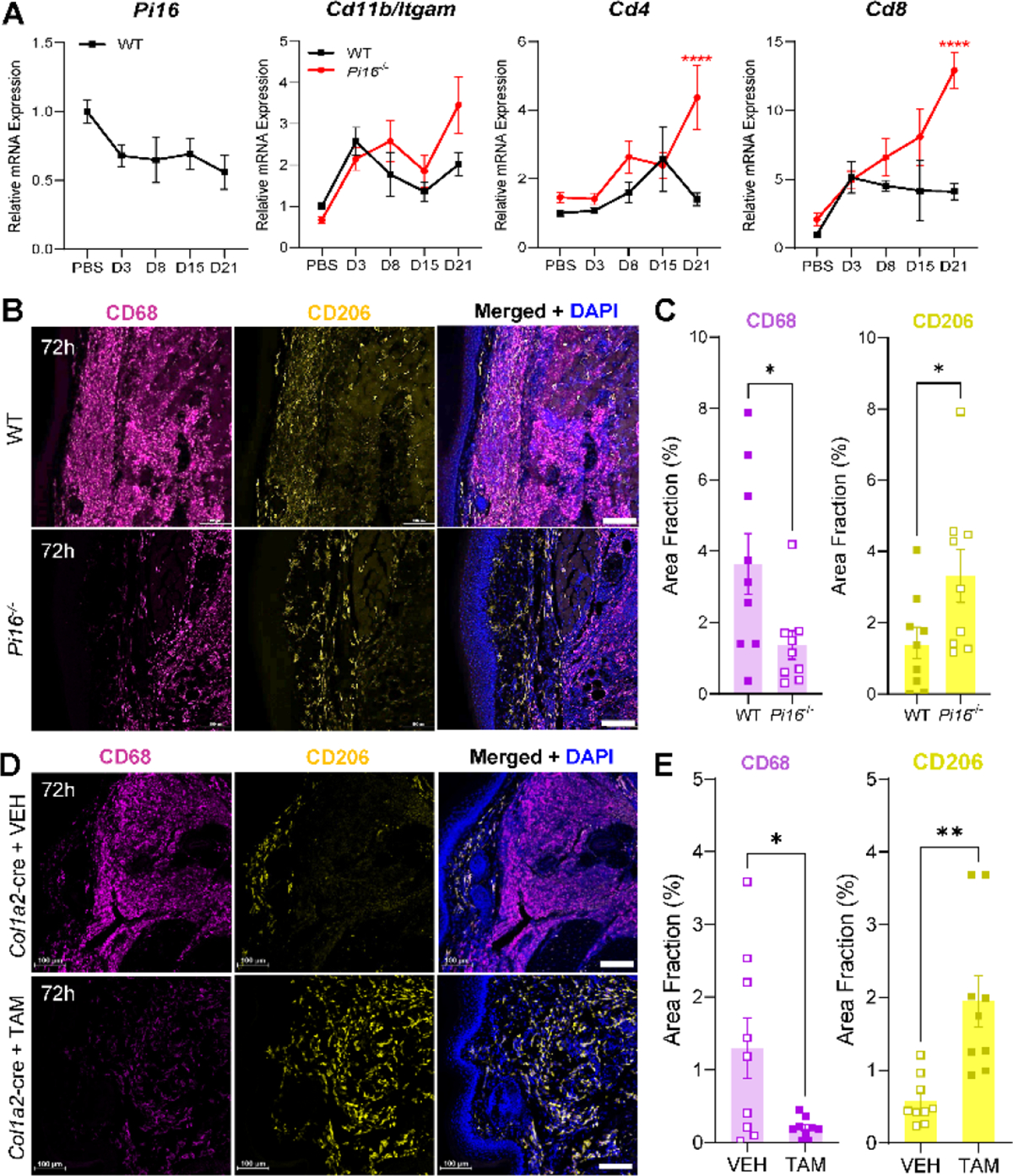
Originally identified in fibroblasts, Protease Inhibitor (PI)16 was recently shown to be crucial for the development of neuropathic pain via effects on blood-nerve barrier permeability and leukocyte infiltration, though its impact on inflammatory pain has not been established. Using the complete Freund's Adjuvant inflammatory pain model, we show that Pi16-/- mice are protected against sustained inflammatory pain. Accordingly, intrathecal delivery of a PI16 neutralizing antibody in wild-type mice prevented sustained CFA pain. In contrast to neuropathic pain models, we did not observe any changes in blood-nerve barrier permeability due to PI16 deletion. Instead, Pi16-/- mice display reduced macrophage density in the CFA-injected hindpaw. Furthermore, there was a significant bias toward CD206hi (anti-inflammatory) macrophages in the hindpaw and associated dorsal root ganglia. Following CFA, intrathecal depletion of CD206+ macrophages using mannosylated clodronate liposomes promoted sustained pain in Pi16-/- mice. Similarly, an IL-10 neutralizing antibody also promoted sustained CFA pain in the Pi16-/ when administered intrathecally. Collectively, our results point to fibroblast-derived PI16 mediating substantial differences in macrophage phenotype in the pain neuroaxis under conditions of inflammation. The co-expression of PI16 alongside fibroblast markers in human DRG raise the likelihood that a similar mechanism operates in human inflammatory pain states. Collectively, our findings may have implications for targeting fibroblast-immune cell crosstalk for the treatment of chronic pain.
Keywords: Blood-nerve barrier; CD206; Complete Freund’s Adjuvant; Fibroblast; IL-10; Inflammation; Macrophage; Meninges; PI16; Pain.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
Comment in
-
Macrophage is more than a bystander in peripheral sensitisation.Brain Behav Immun. 2024 Mar;117:310-311. doi: 10.1016/j.bbi.2024.01.223. Epub 2024 Feb 1. Brain Behav Immun. 2024. PMID: 38307444 No abstract available.
References
-
- Andrews S (2010). “A quality control tool for high throughput sequence data.”, from http://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
-
- Bayik D, Bartels CF, Lovrenert K, Watson DC, Zhang D, Kay K, Lee J, Lauko A, Johnson S, Lo A, Silver DJ, McGraw M, Grabowski M, Mohammadi AM, Veglia F, Fan Y, Vogelbaum MA, Scacheri P and Lathia JD (2022). “Distinct Cell Adhesion Signature Defines Glioblastoma Myeloid-Derived Suppressor Cell Subsets.” Cancer Res 82(22): 4274–4287. - PMC - PubMed
-
- Becker E, Dedden M, Gall C, Wiendl M, Ekici AB, Schulz-Kuhnt A, Schweda A, Voskens C, Hegazy A, Vitali F, Atreya R, Müller TM, Atreya I, Neurath MF and Zundler S (2021). “Residual homing of α4β7-expressing β1(+)PI16(+) regulatory T cells with potent suppressive activity correlates with exposure-efficacy of vedolizumab.” Gut. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
