

A 14-day pulse of PLX5622 modifies α-synucleinopathy in preformed fibril-infused aged mice of both sexes
- PMID: 37315905
- PMCID: PMC10528721
- DOI: 10.1016/j.nbd.2023.106196
A 14-day pulse of PLX5622 modifies α-synucleinopathy in preformed fibril-infused aged mice of both sexes
Abstract
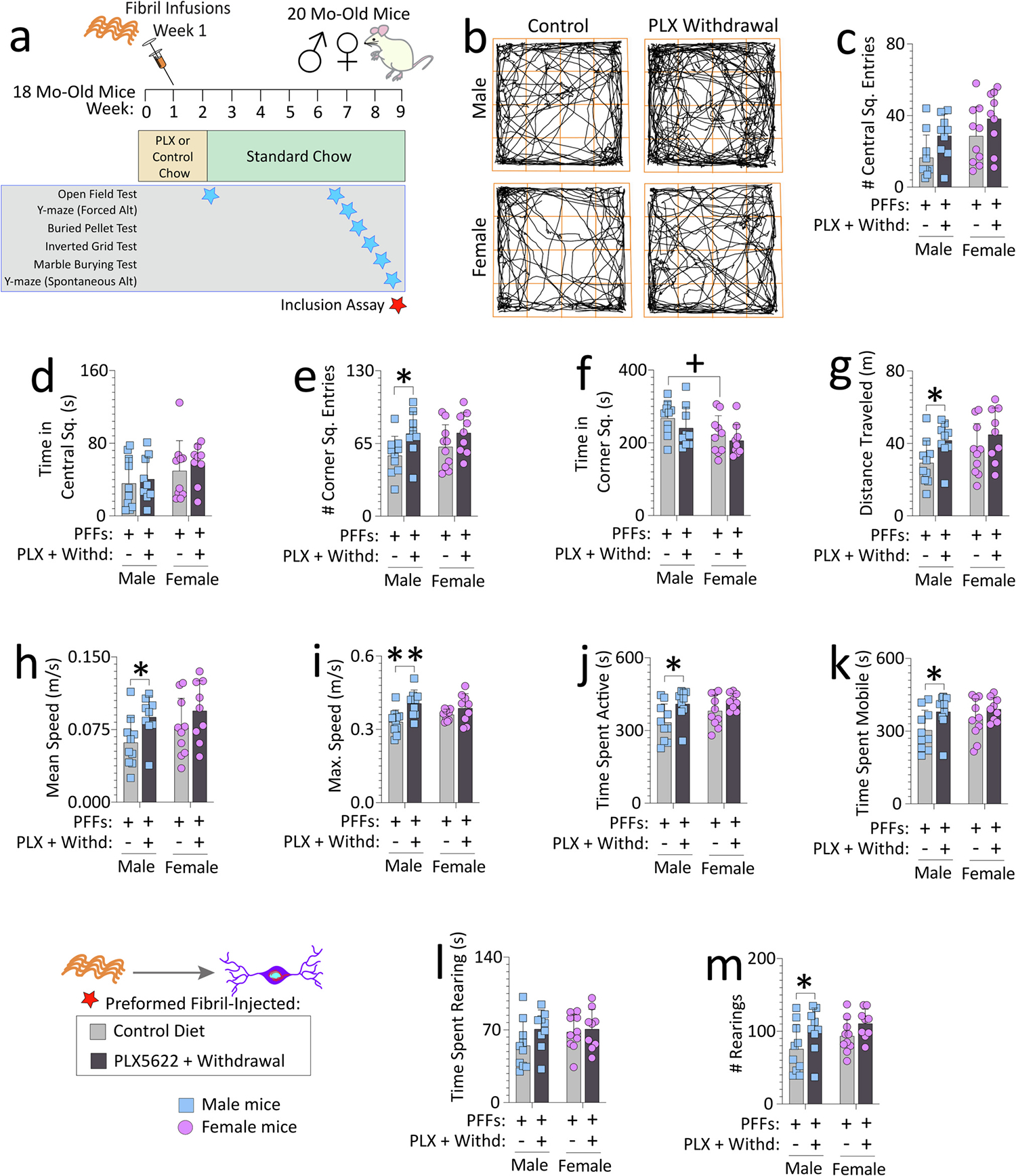
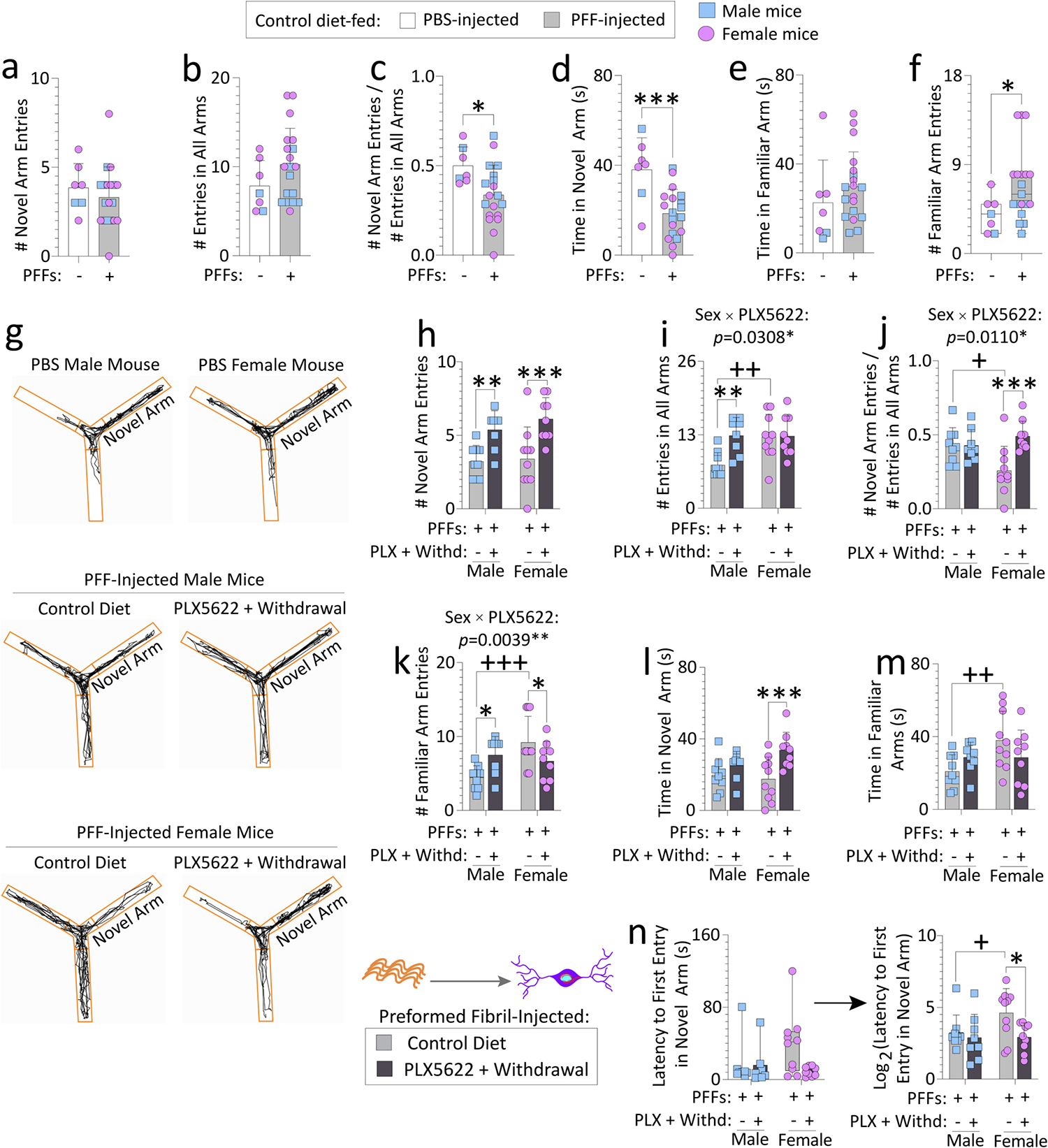
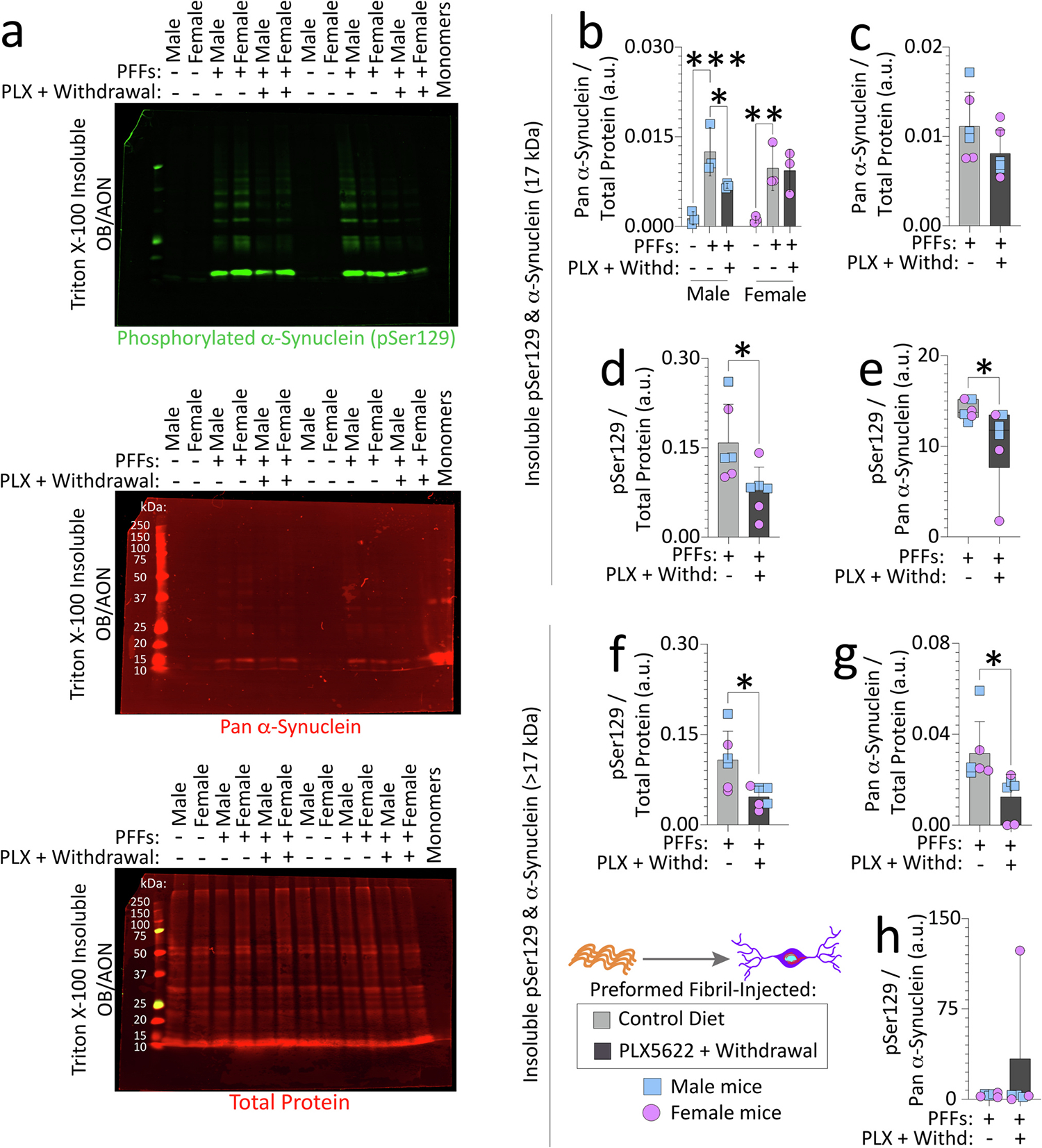
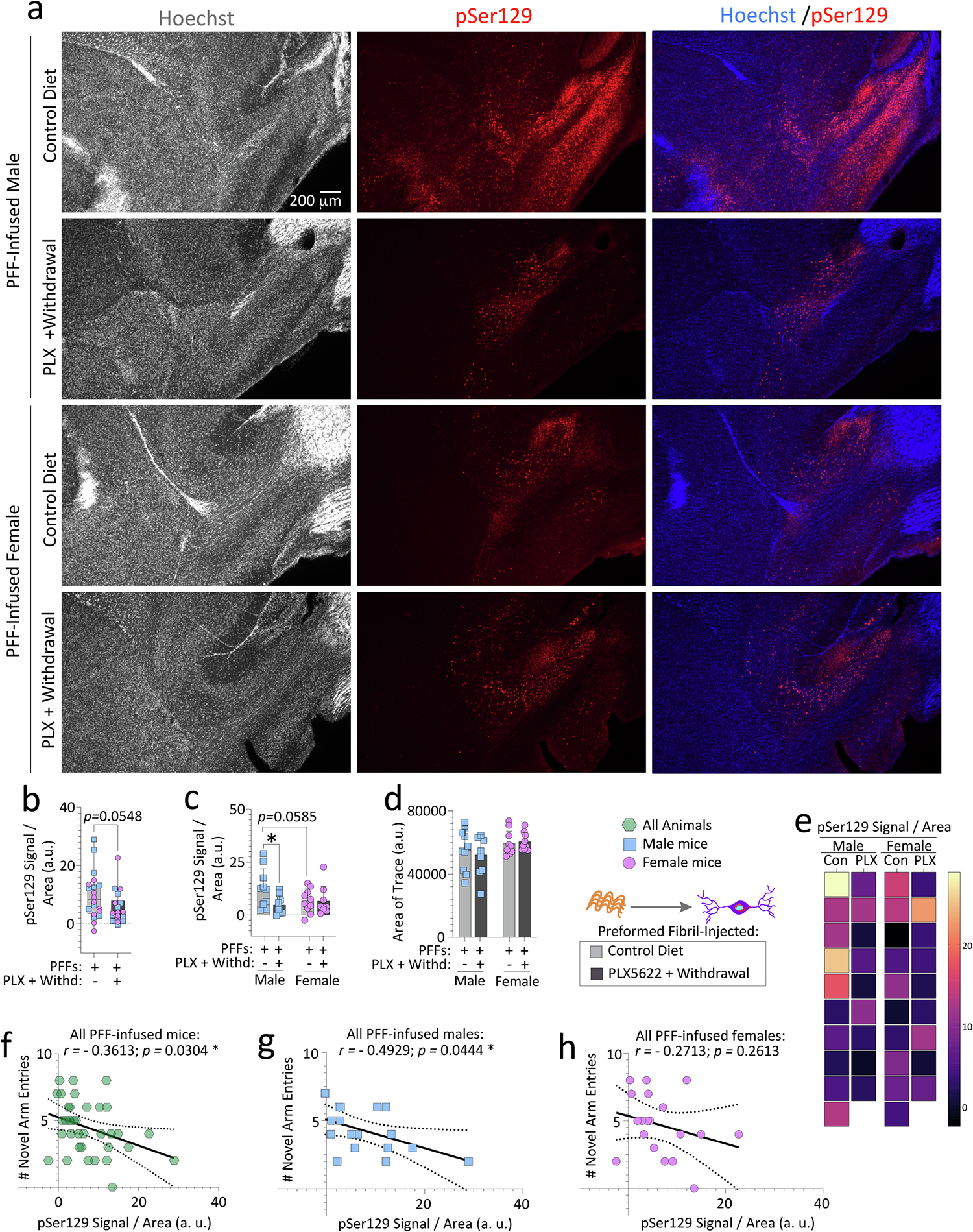
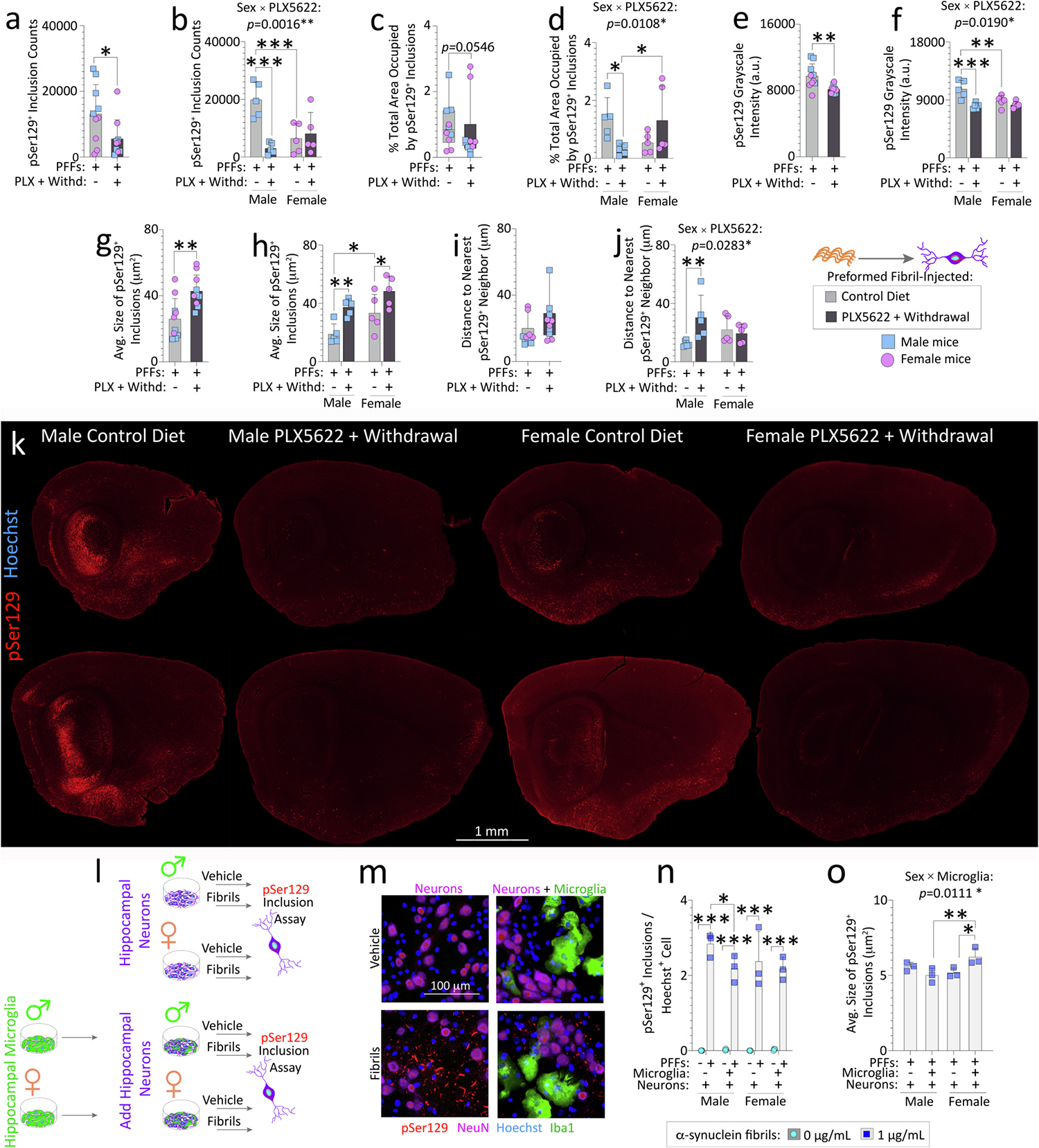
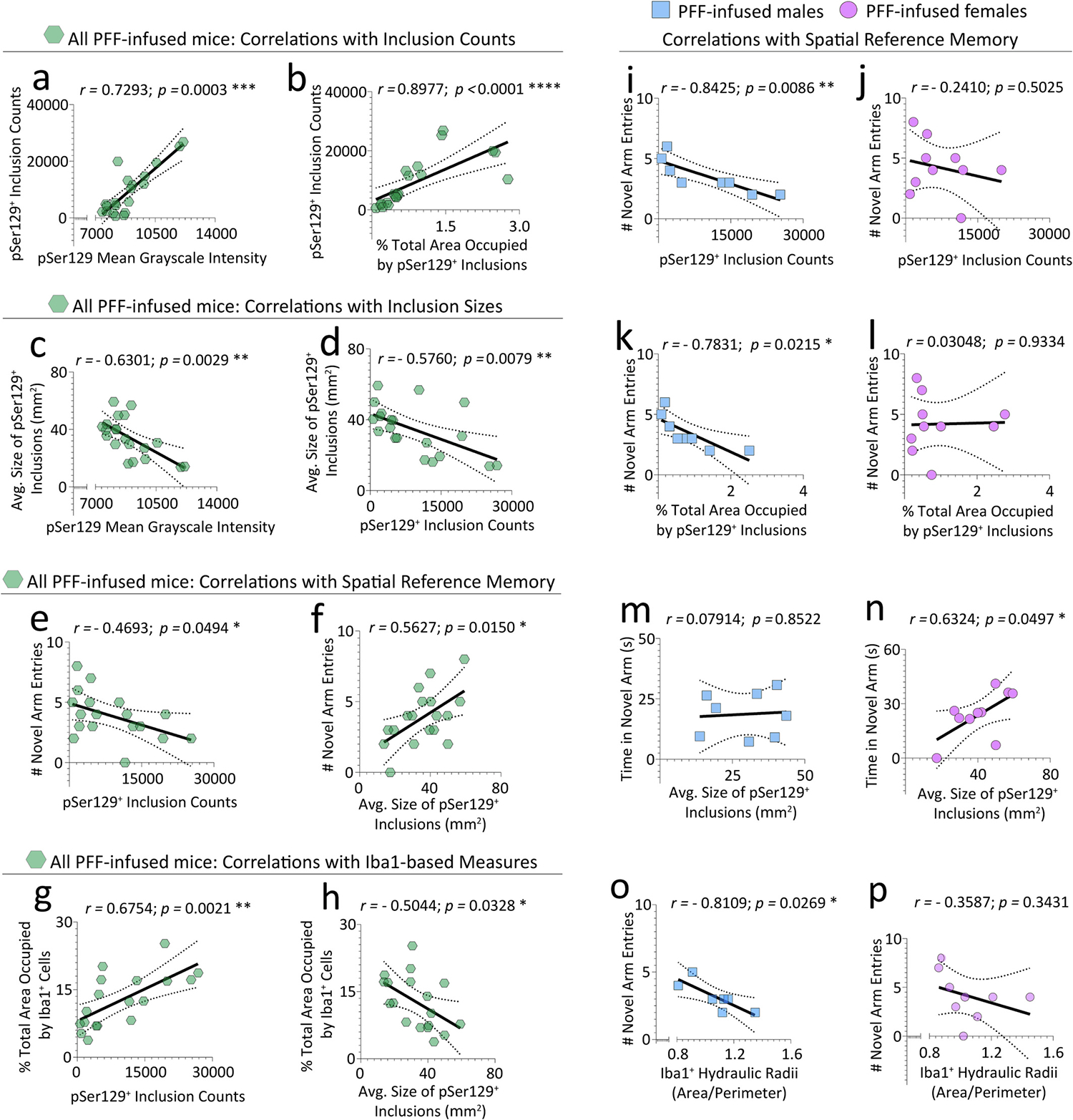
Reactive microglia are observed with aging and in Lewy body disorders, including within the olfactory bulb of men with Parkinson's disease. However, the functional impact of microglia in these disorders is still debated. Resetting these reactive cells by a brief dietary pulse of the colony-stimulating factor 1 receptor (CSF1R) inhibitor PLX5622 may hold therapeutic potential against Lewy-related pathologies. To our knowledge, withdrawal of PLX5622 after short-term exposure has not been tested in the preformed α-synuclein fibril (PFF) model, including in aged mice of both sexes. Compared to aged female mice, we report that aged males on the control diet showed higher numbers of phosphorylated α-synuclein+ inclusions in the limbic rhinencephalon after PFFs were injected in the posterior olfactory bulb. However, aged females displayed larger inclusion sizes compared to males. Short-term (14-day) dietary exposure to PLX5622 followed by control chow reduced inclusion numbers and levels of insoluble α-synuclein in aged males-but not females-and unexpectedly raised inclusion sizes in both sexes. Transient delivery of PLX5622 also improved spatial reference memory in PFF-infused aged mice, as evidenced by an increase in novel arm entries in a Y-maze. Superior memory was positively correlated with inclusion sizes but negatively correlated with inclusion numbers. Although we caution that PLX5622 delivery must be tested further in models of α-synucleinopathy, our data suggest that larger-sized-but fewer-α-synucleinopathic structures are associated with better neurological outcomes in PFF-infused aged mice.
Keywords: Dementia; Lewy body; Microglia; Neurodegeneration; Olfactory bulb; PLX5622; Parkinson's disease; Preformed fibril; Sex; Synuclein.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors have no competing interests to declare.
Figures
Similar articles
-
The center of olfactory bulb-seeded α-synucleinopathy is the limbic system and the ensuing pathology is higher in male than in female mice.Brain Pathol. 2019 Nov;29(6):741-770. doi: 10.1111/bpa.12718. Epub 2019 Apr 10. Brain Pathol. 2019. PMID: 30854742 Free PMC article.
-
Critical appraisal of pathology transmission in the α-synuclein fibril model of Lewy body disorders.Exp Neurol. 2018 Jan;299(Pt A):172-196. doi: 10.1016/j.expneurol.2017.10.017. Epub 2017 Oct 19. Exp Neurol. 2018. PMID: 29056362 Free PMC article.
-
α-synucleinopathy exerts sex-dimorphic effects on the multipurpose DNA repair/redox protein APE1 in mice and humans.Prog Neurobiol. 2022 Sep;216:102307. doi: 10.1016/j.pneurobio.2022.102307. Epub 2022 Jun 13. Prog Neurobiol. 2022. PMID: 35710046 Free PMC article.
-
A Combined α-Synuclein/Fibril (SynFib) Model of Parkinson-Like Synucleinopathy Targeting the Nigrostriatal Dopamine System.J Parkinsons Dis. 2022;12(8):2307-2320. doi: 10.3233/JPD-223452. J Parkinsons Dis. 2022. PMID: 36189605 Free PMC article. Review.
-
Propagation of alpha-synuclein pathology from the olfactory bulb: possible role in the pathogenesis of dementia with Lewy bodies.Cell Tissue Res. 2018 Jul;373(1):233-243. doi: 10.1007/s00441-017-2733-6. Epub 2017 Dec 1. Cell Tissue Res. 2018. PMID: 29196808 Review.
Cited by
-
Altered Cerebrospinal Fluid Proteins in Smith-Lemli-Opitz Syndrome.J Proteome Res. 2025 Aug 1;24(8):4154-4165. doi: 10.1021/acs.jproteome.5c00282. Epub 2025 Jul 9. J Proteome Res. 2025. PMID: 40631482 Free PMC article.
-
Current insights and assumptions on α-synuclein in Lewy body disease.Acta Neuropathol. 2024 Aug 14;148(1):18. doi: 10.1007/s00401-024-02781-3. Acta Neuropathol. 2024. PMID: 39141121 Free PMC article. Review.
-
The variance in phosphorylated, insoluble ⍺-synuclein in humans, rats, and mice is not mainly driven by biological sex.Acta Neuropathol. 2023 Oct;146(4):651-654. doi: 10.1007/s00401-023-02621-w. Epub 2023 Aug 11. Acta Neuropathol. 2023. PMID: 37568067 Free PMC article. No abstract available.
-
Microglia depletion reduces neurodegeneration and remodels extracellular matrix in a mouse Parkinson's disease model triggered by α-synuclein overexpression.NPJ Parkinsons Dis. 2025 Jan 9;11(1):15. doi: 10.1038/s41531-024-00846-4. NPJ Parkinsons Dis. 2025. PMID: 39779738 Free PMC article.
References
-
- Anderson JP, Walker DE, Goldstein JM, de Laat R, Banducci K, Caccavello RJ, Barbour R, Huang J, Kling K, Lee M, Diep L, Keim PS, Shen X, Chataway T, Schlossmacher MG, Seubert P, Schenk D, Sinha S, Gai WP, Chilcote TJ, 2006. Phosphorylation of Ser-129 is the dominant pathological modification of alpha-synuclein in familial and sporadic Lewy body disease. J. Biol. Chem. 281, 29739–29752. - PubMed
-
- Angelova DM, Brown DR, 2019. Microglia and the aging brain: are senescent microglia the key to neurodegeneration? J. Neurochem. 151, 676–688. - PubMed
-
- Attems J, Walker L, Jellinger KA, 2014. Olfactory bulb involvement in neurodegenerative diseases. Acta Neuropathol. 127, 459–475. - PubMed
-
- Attems J, Toledo JB, Walker L, Gelpi E, Gentleman S, Halliday G, Hortobagyi T, Jellinger K, Kovacs GG, Lee EB, Love S, McAleese KE, Nelson PT, Neumann M, Parkkinen L, Polvikoski T, Sikorska B, Smith C, Grinberg LT, Thal DR, Trojanowski JQ, McKeith IG, 2021. Neuropathological consensus criteria for the evaluation of Lewy pathology in post-mortem brains: a multi-Centre study. Acta Neuropathol. 141, 159–172. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
