

Metabolic engineering of Halomonas campaniensis strain XH26 to remove competing pathways to enhance ectoine production
- PMID: 37322079
- PMCID: PMC10272175
- DOI: 10.1038/s41598-023-36975-8
Metabolic engineering of Halomonas campaniensis strain XH26 to remove competing pathways to enhance ectoine production
Abstract
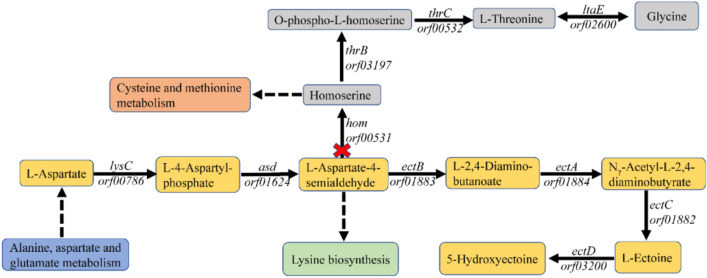
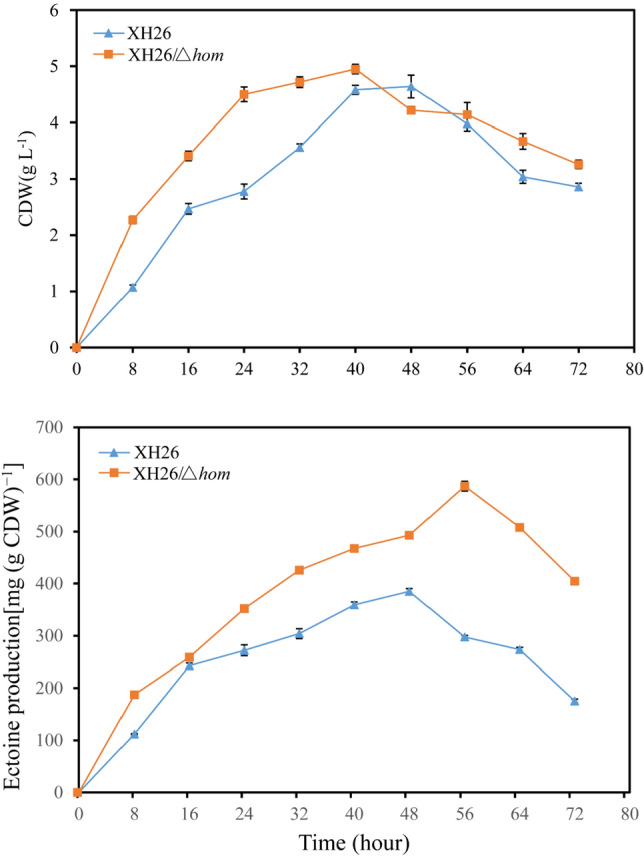
Ectoine has gained considerable attention as a high-value chemical with significant application potential and market demand. This study aimed to increase ectoine yields by blocking the metabolic shunt pathway of L-aspartate-4-semialdehyde, the precursor substrate in ectoine synthesis. The homoserine dehydrogenase encoded by hom in H. campaniensis strain XH26 is responsible for the metabolic shunt of L-aspartate-4-semialdehyde to glycine. CRISPR/Cas9 technology was used to seamlessly knockout hom, blocking the metabolic shunt pathway to increase ectoine yields. The ectoine yield of XH26/Δhom was 351.13 mg (g CDW)-1 after 48 h of incubation in 500 mL shake flasks using optimal medium with 1.5 mol L-1 NaCl, which was significantly higher than the 239.18 mg (g CDW)-1 of the wild-type strain. Additionally, the absence of the ectoine metabolic shunt pathway affects betaine synthesis, and thus the betaine yields of XH26/Δhom was 19.98 mg (g CDW)-1, considerably lower than the 69.58 mg (g CDW)-1 of the wild-type strain. Batch fermentation parameters were optimized, and the wild-type strain and XH26/Δhom were fermented in 3 L fermenters, resulting in a high ectoine yield of 587.09 mg (g CDW)-1 for the defective strain, which was significantly greater than the ectoine yield of 385.03 mg (g CDW)-1 of the wild-type strain. This study showed that blocking the metabolic shunt of synthetic substrates effectively increases ectoine production, and a reduction in the competitively compatible solute betaine appears to promote increased ectoine synthesis.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Comparative genomic analysis of Halomonas campaniensis wild-type and ultraviolet radiation-mutated strains reveal genomic differences associated with increased ectoine production.Int Microbiol. 2023 Nov;26(4):1009-1020. doi: 10.1007/s10123-023-00356-y. Epub 2023 Apr 17. Int Microbiol. 2023. PMID: 37067733 Free PMC article.
-
Study of osmoadaptation mechanisms of halophilic Halomonas alkaliphila XH26 under salt stress by transcriptome and ectoine analysis.Extremophiles. 2022 Mar 1;26(1):14. doi: 10.1007/s00792-022-01256-1. Extremophiles. 2022. PMID: 35229247
-
High Ectoine Production from Lignocellulosic Hydrolysate by Escherichia coli through Metabolic and Fermentation Engineering.ACS Synth Biol. 2025 Feb 21;14(2):609-620. doi: 10.1021/acssynbio.4c00899. Epub 2025 Feb 11. ACS Synth Biol. 2025. PMID: 39933098
-
Microbial Production of Ectoine: A Review.ACS Synth Biol. 2025 Feb 21;14(2):332-342. doi: 10.1021/acssynbio.4c00490. Epub 2025 Jan 20. ACS Synth Biol. 2025. PMID: 39834017 Review.
-
Microbial production of ectoine and hydroxyectoine as high-value chemicals.Microb Cell Fact. 2021 Mar 26;20(1):76. doi: 10.1186/s12934-021-01567-6. Microb Cell Fact. 2021. PMID: 33771157 Free PMC article. Review.
Cited by
-
Metabolic engineering of Bacillus licheniformis DW2 for ectoine production.World J Microbiol Biotechnol. 2025 Jan 2;41(1):23. doi: 10.1007/s11274-024-04238-x. World J Microbiol Biotechnol. 2025. PMID: 39743645
-
Halomonas flagellata sp. nov., a halophilic bacterium isolated from saline soil in Xinjiang.Arch Microbiol. 2023 Sep 26;205(10):340. doi: 10.1007/s00203-023-03670-1. Arch Microbiol. 2023. PMID: 37750964
-
Establishing Halomonas as a chassis for industrial biotechnology: advances in synthetic biology tool development and metabolic engineering strategies.Microb Cell Fact. 2025 Jun 12;24(1):133. doi: 10.1186/s12934-025-02757-2. Microb Cell Fact. 2025. PMID: 40506695 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
