

Activation of the Aryl Hydrocarbon Receptor in Muscle Exacerbates Ischemic Pathology in Chronic Kidney Disease
- PMID: 37325935
- PMCID: PMC10330629
- DOI: 10.1161/CIRCRESAHA.123.322875
Activation of the Aryl Hydrocarbon Receptor in Muscle Exacerbates Ischemic Pathology in Chronic Kidney Disease
Abstract
Background: Chronic kidney disease (CKD) accelerates the development of atherosclerosis, decreases muscle function, and increases the risk of amputation or death in patients with peripheral artery disease (PAD). However, the mechanisms underlying this pathobiology are ill-defined. Recent work has indicated that tryptophan-derived uremic solutes, which are ligands for AHR (aryl hydrocarbon receptor), are associated with limb amputation in PAD. Herein, we examined the role of AHR activation in the myopathy of PAD and CKD.
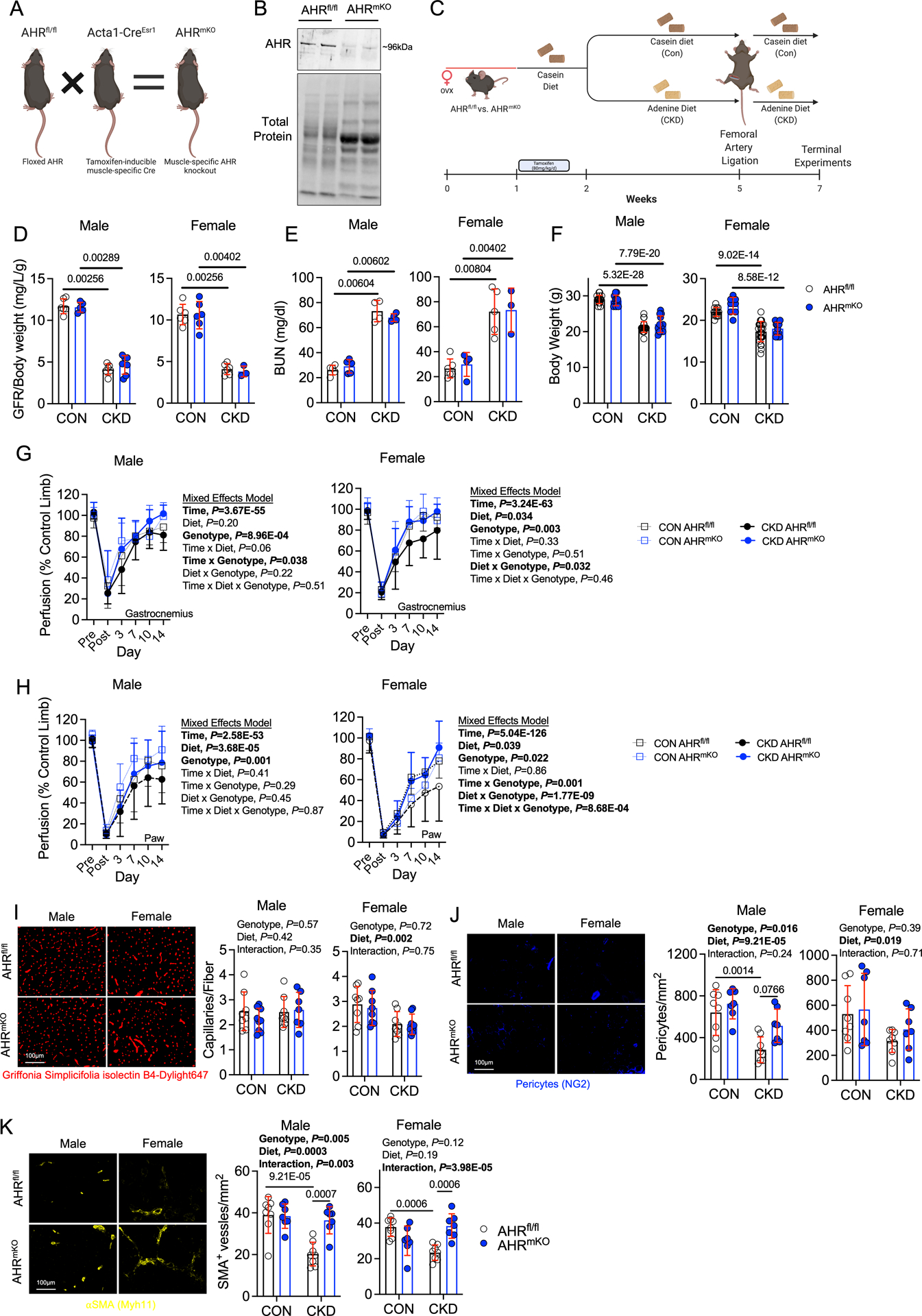
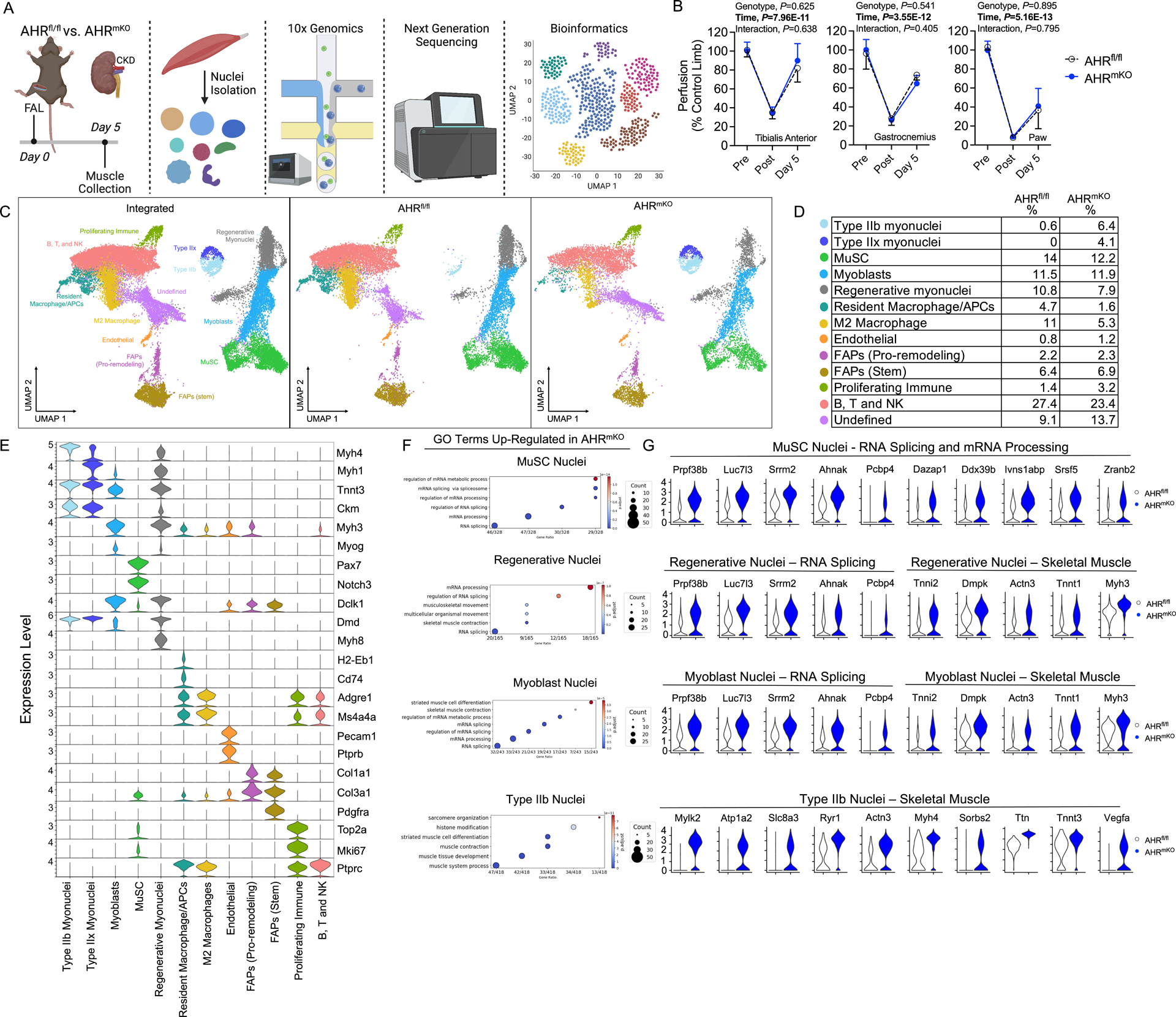
Methods: AHR-related gene expression was evaluated in skeletal muscle obtained from mice and human PAD patients with and without CKD. AHRmKO (skeletal muscle-specific AHR knockout) mice with and without CKD were subjected to femoral artery ligation, and a battery of assessments were performed to evaluate vascular, muscle, and mitochondrial health. Single-nuclei RNA sequencing was performed to explore intercellular communication. Expression of the constitutively active AHR was used to isolate the role of AHR in mice without CKD.
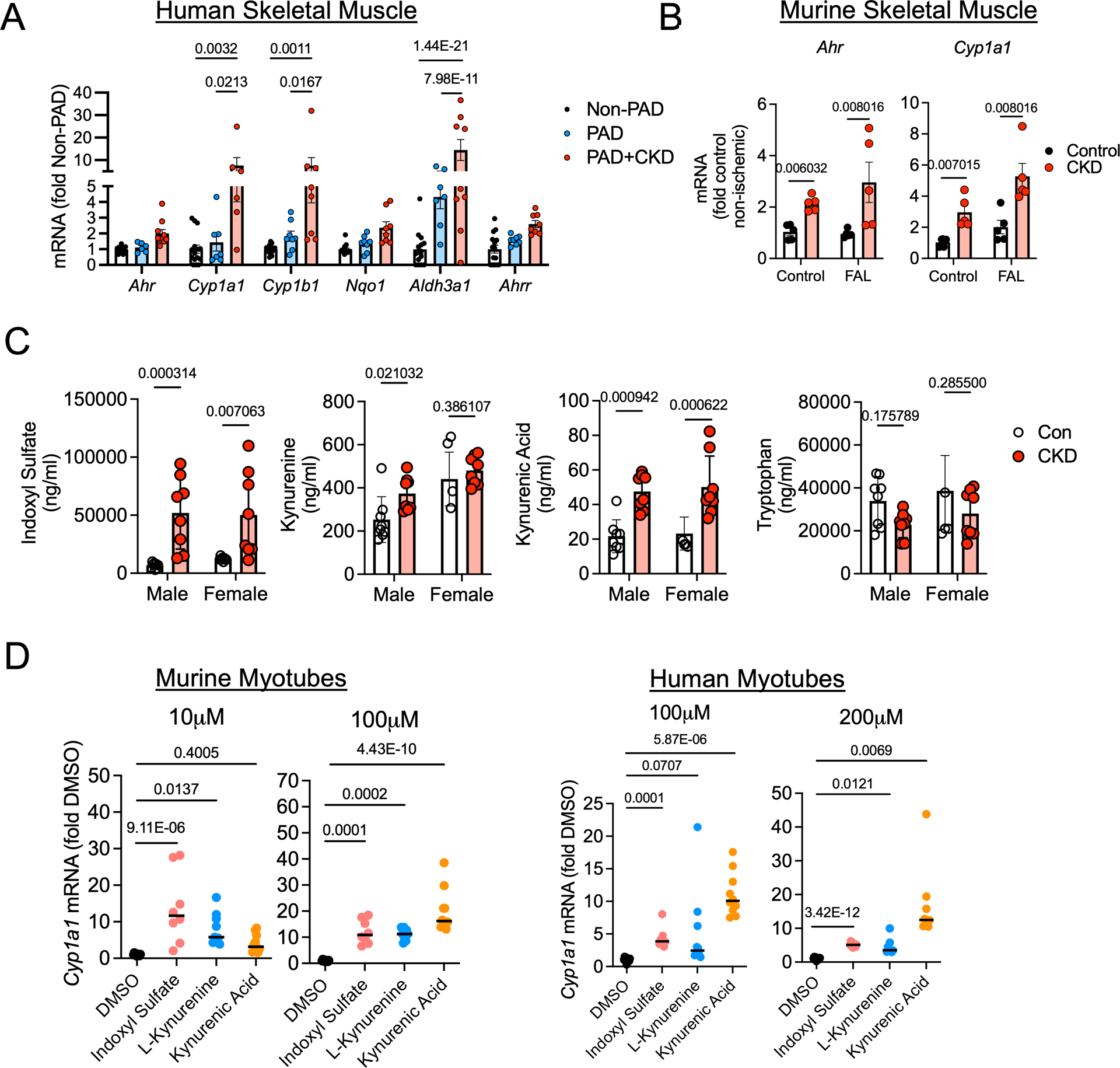
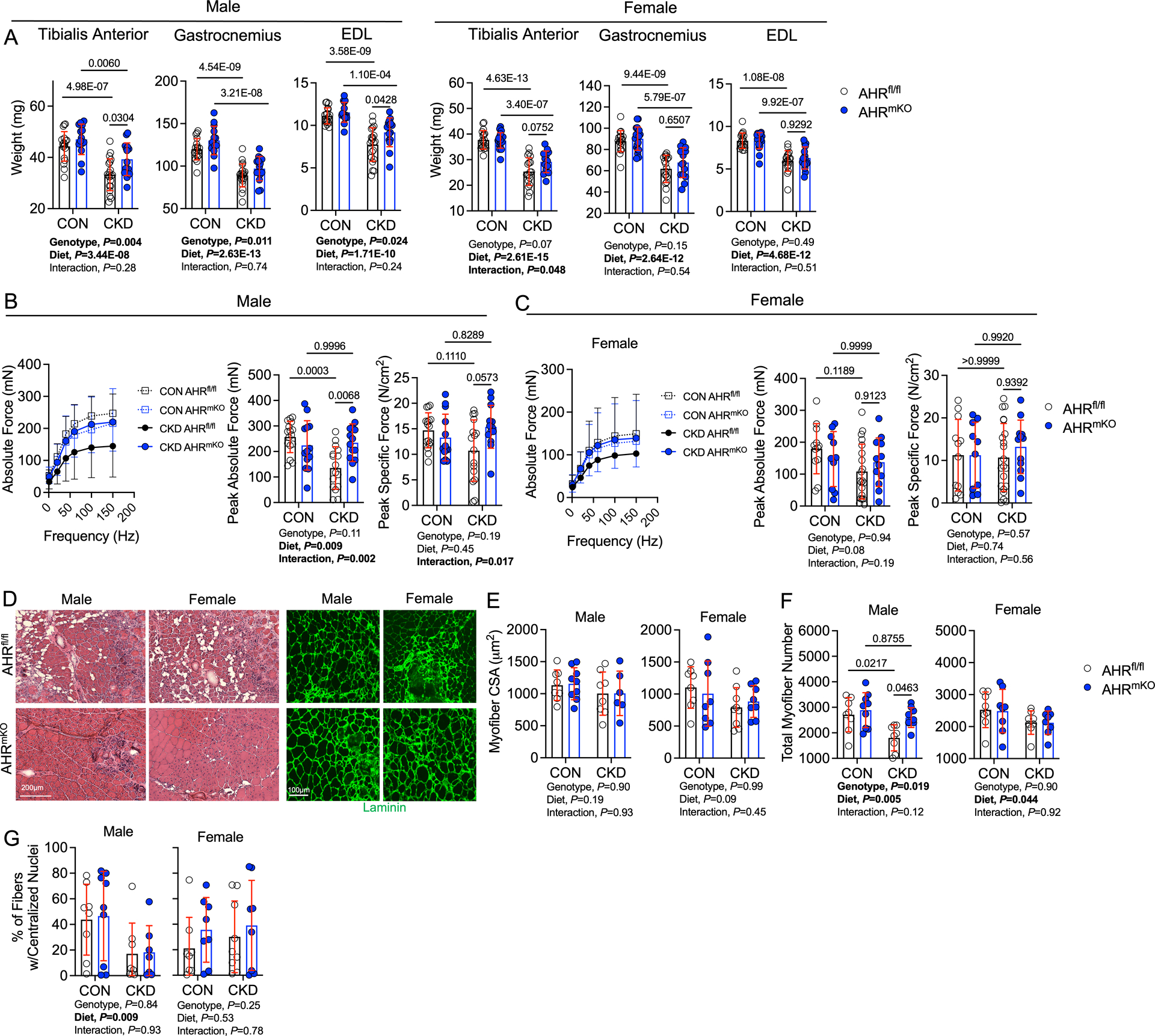
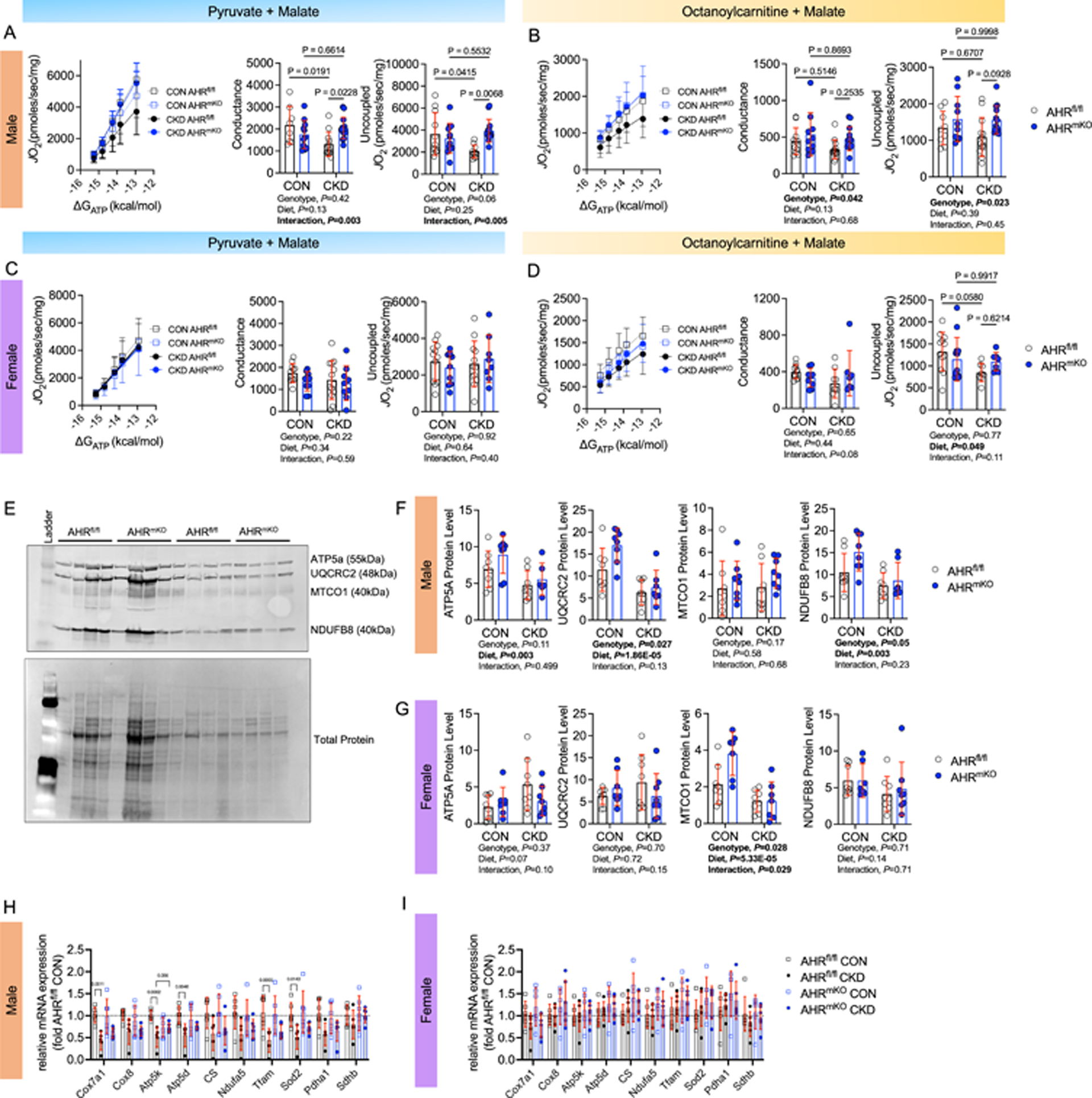
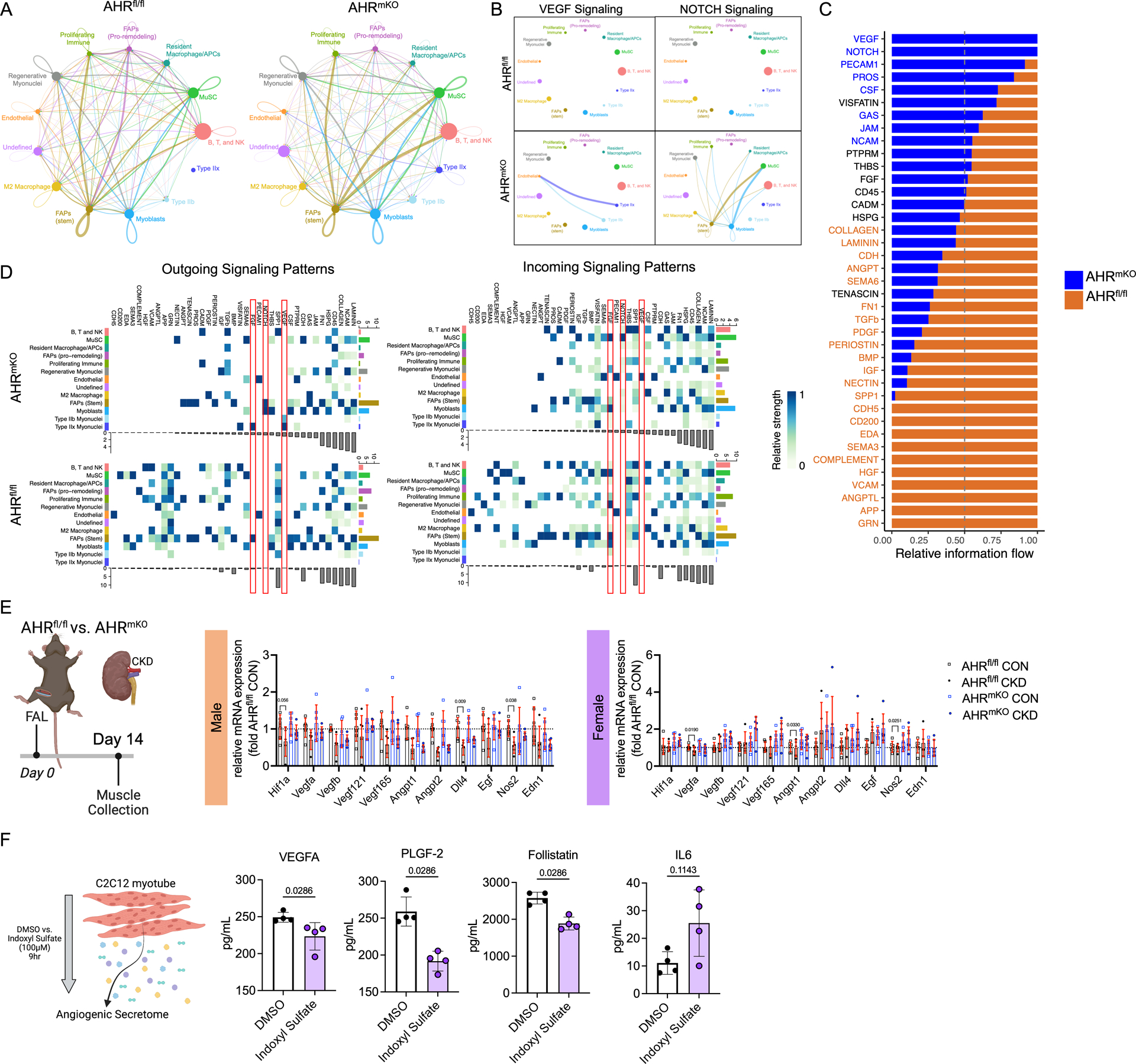
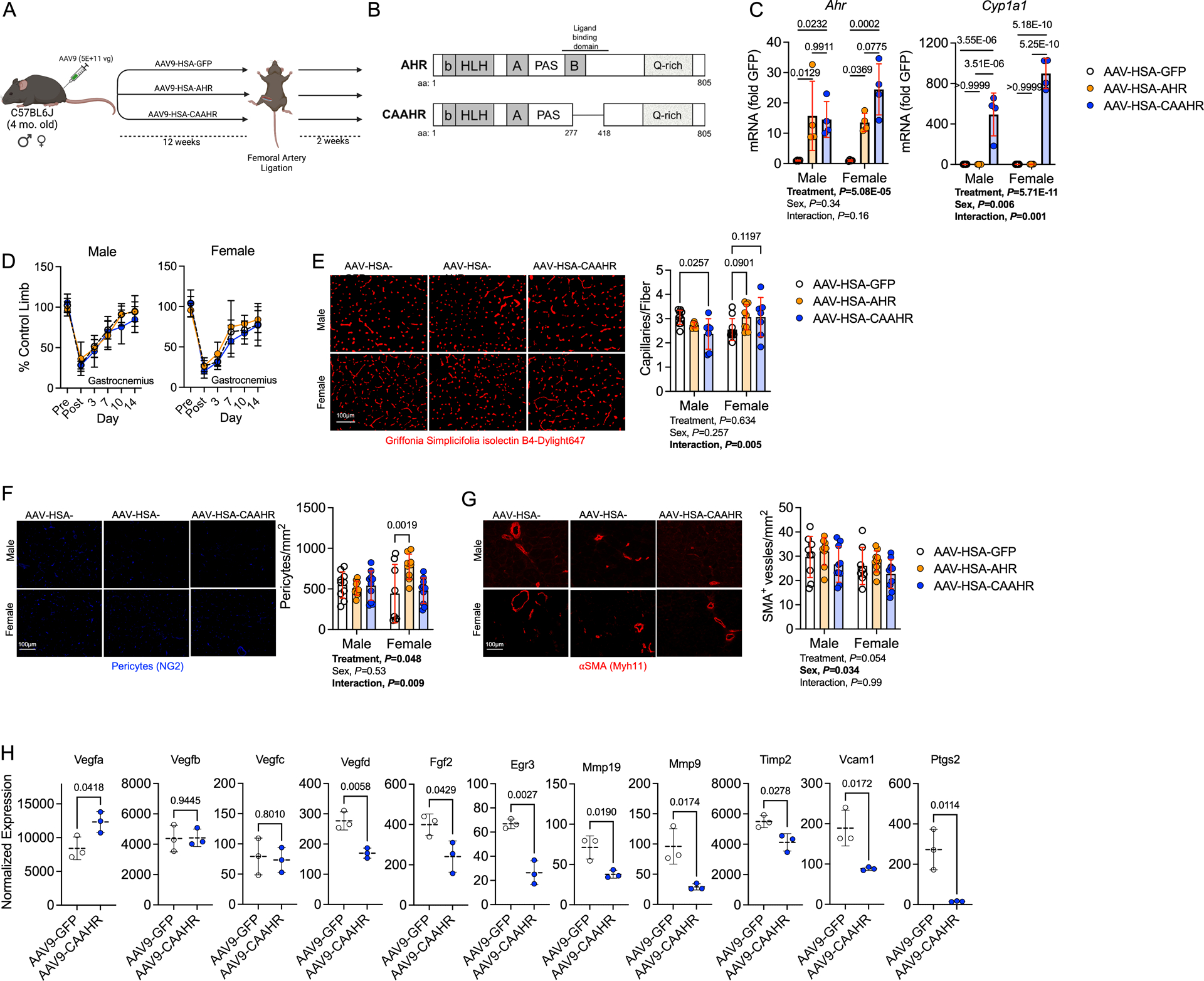
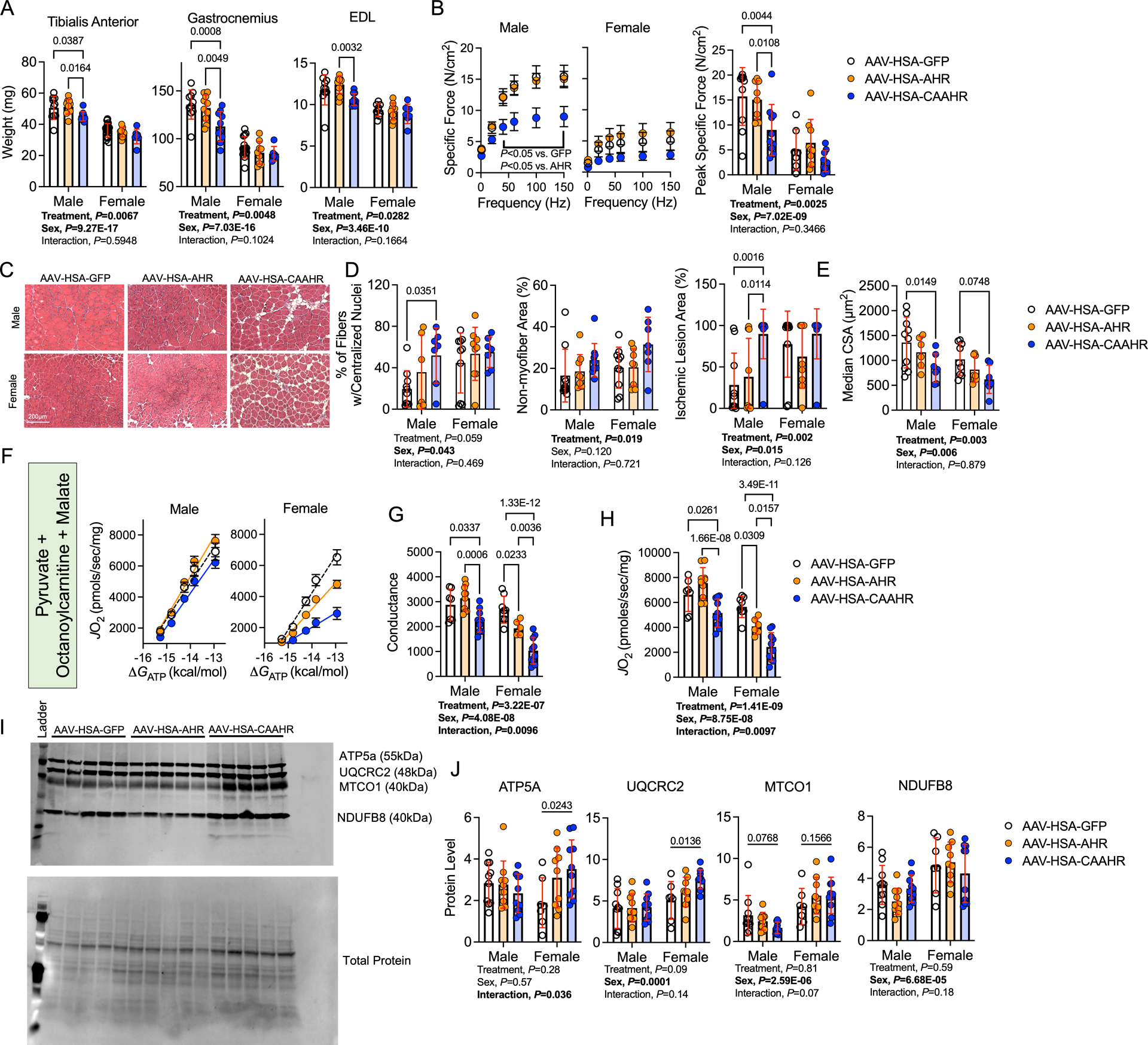
Results: PAD patients and mice with CKD displayed significantly higher mRNA expression of classical AHR-dependent genes (Cyp1a1, Cyp1b1, and Aldh3a1) when compared with either muscle from the PAD condition with normal renal function (P<0.05 for all 3 genes) or nonischemic controls. AHRmKO significantly improved limb perfusion recovery and arteriogenesis, preserved vasculogenic paracrine signaling from myofibers, increased muscle mass and strength, as well as enhanced mitochondrial function in an experimental model of PAD/CKD. Moreover, viral-mediated skeletal muscle-specific expression of a constitutively active AHR in mice with normal kidney function exacerbated the ischemic myopathy evidenced by smaller muscle masses, reduced contractile function, histopathology, altered vasculogenic signaling, and lower mitochondrial respiratory function.
Conclusions: These findings establish AHR activation in muscle as a pivotal regulator of the ischemic limb pathology in CKD. Further, the totality of the results provides support for testing of clinical interventions that diminish AHR signaling in these conditions.
Keywords: humans; kidney; mitochondria; peripheral artery disease; uremia.
Conflict of interest statement
Figures
Update of
-
Chronic activation of the aryl hydrocarbon receptor in muscle exacerbates ischemic pathology in chronic kidney disease.bioRxiv [Preprint]. 2023 May 18:2023.05.16.541060. doi: 10.1101/2023.05.16.541060. bioRxiv. 2023. Update in: Circ Res. 2023 Jul 7;133(2):158-176. doi: 10.1161/CIRCRESAHA.123.322875. PMID: 37292677 Free PMC article. Updated. Preprint.
Similar articles
-
Deletion of the aryl hydrocarbon receptor in endothelial cells improves ischemic angiogenesis in chronic kidney disease.Am J Physiol Heart Circ Physiol. 2024 Jan 1;326(1):H44-H60. doi: 10.1152/ajpheart.00530.2023. Epub 2023 Nov 3. Am J Physiol Heart Circ Physiol. 2024. PMID: 37921663 Free PMC article.
-
Multiomic Analysis of Calf Muscle in Peripheral Artery Disease and Chronic Kidney Disease.Circ Res. 2025 Mar 28;136(7):688-703. doi: 10.1161/CIRCRESAHA.124.325642. Epub 2025 Feb 18. Circ Res. 2025. PMID: 39963788 Free PMC article.
-
Chronic activation of the aryl hydrocarbon receptor in muscle exacerbates ischemic pathology in chronic kidney disease.bioRxiv [Preprint]. 2023 May 18:2023.05.16.541060. doi: 10.1101/2023.05.16.541060. bioRxiv. 2023. Update in: Circ Res. 2023 Jul 7;133(2):158-176. doi: 10.1161/CIRCRESAHA.123.322875. PMID: 37292677 Free PMC article. Updated. Preprint.
-
The aryl hydrocarbon receptor-activating effect of uremic toxins from tryptophan metabolism: a new concept to understand cardiovascular complications of chronic kidney disease.Toxins (Basel). 2014 Mar 4;6(3):934-49. doi: 10.3390/toxins6030934. Toxins (Basel). 2014. PMID: 24599232 Free PMC article. Review.
-
Nuclear receptors as potential therapeutic targets in peripheral arterial disease and related myopathy.FEBS J. 2023 Oct;290(19):4596-4613. doi: 10.1111/febs.16593. Epub 2022 Aug 18. FEBS J. 2023. PMID: 35942640 Free PMC article. Review.
Cited by
-
The dichotomic role of cytokines in aging.Biogerontology. 2024 Dec 2;26(1):17. doi: 10.1007/s10522-024-10152-4. Biogerontology. 2024. PMID: 39621124 Review.
-
Deletion of the aryl hydrocarbon receptor in endothelial cells improves ischemic angiogenesis in chronic kidney disease.Am J Physiol Heart Circ Physiol. 2024 Jan 1;326(1):H44-H60. doi: 10.1152/ajpheart.00530.2023. Epub 2023 Nov 3. Am J Physiol Heart Circ Physiol. 2024. PMID: 37921663 Free PMC article.
-
Multiomic Analysis of Calf Muscle in Peripheral Artery Disease and Chronic Kidney Disease.Circ Res. 2025 Mar 28;136(7):688-703. doi: 10.1161/CIRCRESAHA.124.325642. Epub 2025 Feb 18. Circ Res. 2025. PMID: 39963788 Free PMC article.
-
Mitochondrial targeted catalase improves muscle strength following arteriovenous fistula creation in mice with chronic kidney disease.Sci Rep. 2024 Apr 9;14(1):8288. doi: 10.1038/s41598-024-58805-1. Sci Rep. 2024. PMID: 38594299 Free PMC article.
-
A tryptophan-derived uremic metabolite/Ahr/Pdk4 axis governs skeletal muscle mitochondrial energetics in chronic kidney disease.JCI Insight. 2024 Apr 23;9(10):e178372. doi: 10.1172/jci.insight.178372. JCI Insight. 2024. PMID: 38652558 Free PMC article.
References
-
- Ostchega Y, Paulose-Ram R, Dillon CF, Gu Q, Hughes JP. Prevalence of peripheral arterial disease and risk factors in persons aged 60 and older: data from the National Health and Nutrition Examination Survey 1999–2004. J Am Geriatr Soc. 2007;55:583–589. doi: 10.1111/j.1532-5415.2007.01123.x - DOI - PubMed
-
- Fowkes FG, Rudan D, Rudan I, Aboyans V, Denenberg JO, McDermott MM, Norman PE, Sampson UK, Williams LJ, Mensah GA, et al. Comparison of global estimates of prevalence and risk factors for peripheral artery disease in 2000 and 2010: a systematic review and analysis. Lancet. 2013;382:1329–1340. doi: 10.1016/S0140-6736(13)61249-0 - DOI - PubMed
-
- Heideman PP, Rajebi MR, McKusick MA, Bjarnason H, Oderich GS, Friese JL, Fleming MD, Stockland AH, Harmsen WS, Mandrekar J, et al. Impact of Chronic Kidney Disease on Clinical Outcomes of Endovascular Treatment for Femoropopliteal Arterial Disease. J Vasc Interv Radiol. 2016;27:1204–1214. doi: 10.1016/j.jvir.2016.04.036 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
