

Sexual selection for both diversity and repetition in birdsong
- PMID: 37328501
- PMCID: PMC10275917
- DOI: 10.1038/s41467-023-39308-5
Sexual selection for both diversity and repetition in birdsong
Abstract
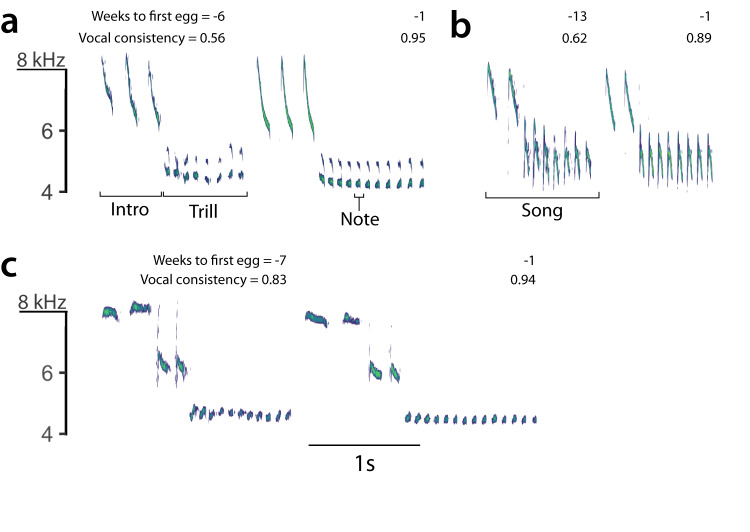
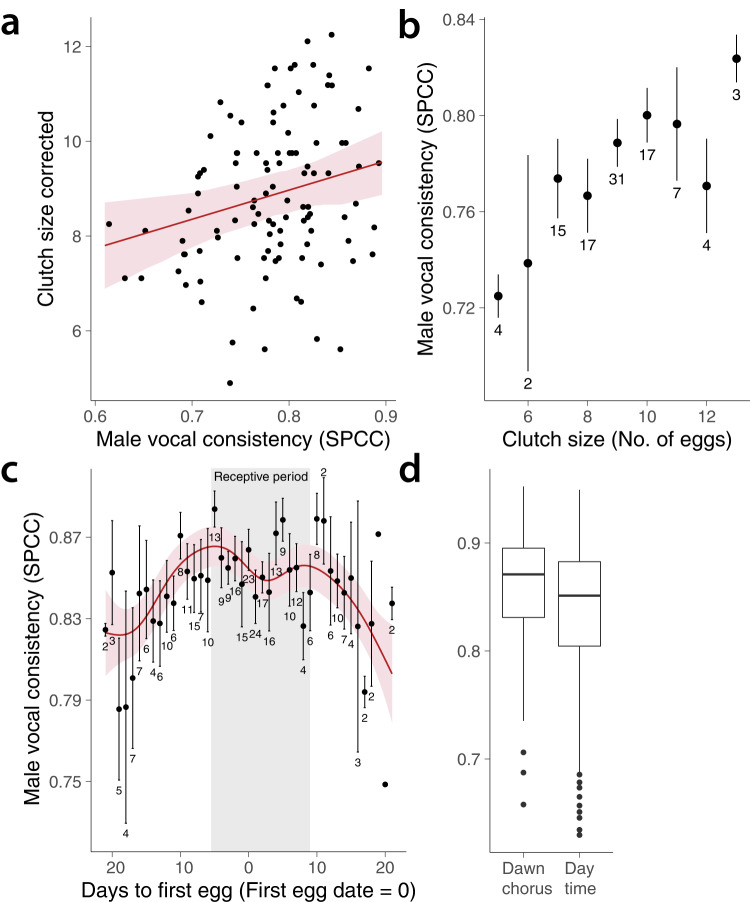
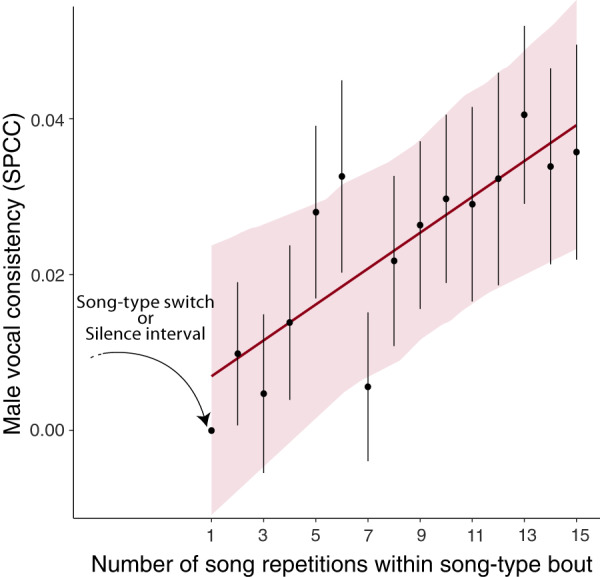
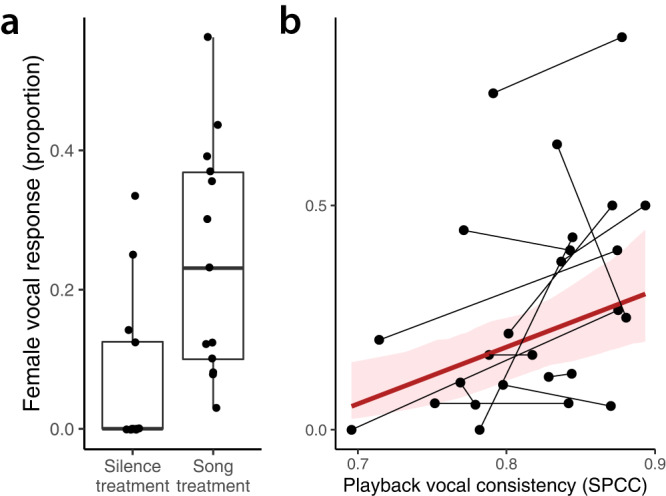
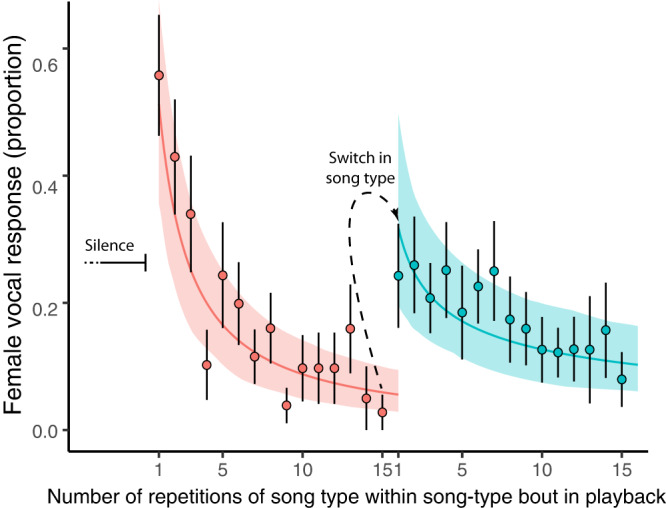
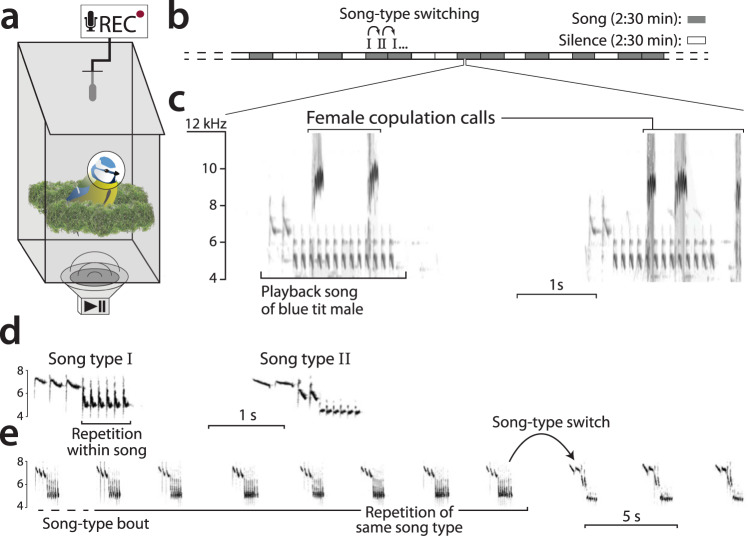
From fiddler crabs to humans, animals perform repetitive displays showing neuromotor skill and vigour. Consistent repetition of identical notes (vocal consistency) facilitates the assessment of neuromotor skills and is important in communication in birds. Most birdsong research has focused on song diversity as a signal of individual quality, which seems contradictory as repetition is extremely common in most species. Here we show that consistent repetition within songs is positively correlated with reproductive success in male blue tits (Cyanistes caeruleus). A playback experiment shows that females are sexually aroused by male songs with high levels of vocal consistency, which also peaks seasonally during the fertile period of the female, supporting the role of vocal consistency in mate choice. Male vocal consistency also increases with subsequent repetitions of the same song type (a warm-up effect) which conflicts with the fact that females habituate to repeated song, showing decreased arousal. Importantly, we find that switching song types elicits significant dishabituation within the playback, supporting the habituation hypothesis as an evolutionary mechanism driving song diversity in birds. An optimal balance between repetition and diversity may explain the singing style of many bird species and displays of other animals.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Female blue tits sing frequently: a sex comparison of occurrence, context, and structure of song.Behav Ecol. 2022 Jun 20;33(5):912-925. doi: 10.1093/beheco/arac044. eCollection 2022 Sep-Oct. Behav Ecol. 2022. PMID: 36382229 Free PMC article.
-
The singing question: re-conceptualizing birdsong.Biol Rev Camb Philos Soc. 2022 Feb;97(1):326-342. doi: 10.1111/brv.12800. Epub 2021 Oct 5. Biol Rev Camb Philos Soc. 2022. PMID: 34609054
-
Physically challenging song traits, male quality, and reproductive success in house wrens.PLoS One. 2013;8(3):e59208. doi: 10.1371/journal.pone.0059208. Epub 2013 Mar 19. PLoS One. 2013. PMID: 23527137 Free PMC article.
-
Birdsong and singing behavior.Ann N Y Acad Sci. 2004 Jun;1016:1-30. doi: 10.1196/annals.1298.029. Ann N Y Acad Sci. 2004. PMID: 15313767 Review.
-
Where to from here? Perspectives on steroid-induced and naturally-occurring singing in female songbirds.Horm Behav. 2022 Feb;138:105098. doi: 10.1016/j.yhbeh.2021.105098. Epub 2021 Dec 10. Horm Behav. 2022. PMID: 34902757 Review.
Cited by
-
A genomic basis of vocal rhythm in birds.Nat Commun. 2024 Apr 23;15(1):3095. doi: 10.1038/s41467-024-47305-5. Nat Commun. 2024. PMID: 38653976 Free PMC article.
-
A limit to sustained performance constrains trill length in birdsong.iScience. 2023 Oct 14;26(11):108206. doi: 10.1016/j.isci.2023.108206. eCollection 2023 Nov 17. iScience. 2023. PMID: 37953962 Free PMC article.
-
Global analysis of acoustic frequency characteristics in birds.Proc Biol Sci. 2024 Nov;291(2034):20241908. doi: 10.1098/rspb.2024.1908. Epub 2024 Nov 6. Proc Biol Sci. 2024. PMID: 39501883
-
Male song structure predicts offspring recruitment to the breeding population in a migratory bird.Evolution. 2024 May 29;78(6):1054-1066. doi: 10.1093/evolut/qpae036. Evolution. 2024. PMID: 38441178 Free PMC article.
-
Adult neurogenesis is necessary for functional regeneration of a forebrain neural circuit.Proc Natl Acad Sci U S A. 2024 Jul 9;121(28):e2400596121. doi: 10.1073/pnas.2400596121. Epub 2024 Jul 5. Proc Natl Acad Sci U S A. 2024. PMID: 38968119 Free PMC article.
References
-
- Bradbury, J. W. & Vehrencamp, S. L. Principles of Animal Communication (Sinauer Associates, 1998).
-
- Byers J, Hebets E, Podos J. Female mate choice based upon male motor performance. Anim. Behav. 2010;79:771–778. doi: 10.1016/j.anbehav.2010.01.009. - DOI
-
- Jordão JM, Curto AF, Oliveira RF. Stereotypy and variation in the claw waving display of the fiddler crab (Uca tangeri) Acta Ethol. 2007;10:55–62. doi: 10.1007/s10211-007-0030-1. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
